Зачатие у человека — это… Что такое Зачатие у человека?
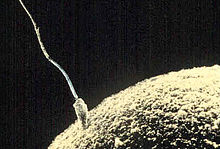
Зачатие (от рус. зачать, то есть начать) — возникновение беременности. Согласно энциклопедическому словарю медицинских терминов зачатие включает оплодотворение (слияние сперматозоида и яйцеклетки), развитие эмбриона до имплантации (дробление) и процессы имплантации (прикрепления эмбриона к слизистой оболочке матки). Таким образом, у человека зачатие охватывает события, происходящие в первые две недели беременности, от полового акта, приведшего к оплодотворению, до появления первых признаков беременности (задержка менструации, повышение уровня хорионического гонадотропина и т.д.)
Оплодотворение и формирование зиготы
При естественном половом акте сперма мужчины попадает во влагалище женщины. Среда влагалища является губительной для сперматозоидов в силу повышенной кислотности (pH около 4). Спустя два часа после эякуляции большая часть сперматозоидов погибает во влагалище, но часть подвижных сперматозоидов проникает в шейку матки и далее в матку. Шейка матки заполнена шеечной слизью, которая является барьером для проникновения микроорганизмов из влагалища в матку, но также шеечная слизь может оказывать препятствие при проникновении сперматозоидов в матку. В патологических случаях чрезмерно вязкая шеечная слизь не позволяет пройти достаточному количеству сперматозоидов в матку, что может явиться причиной бесплодия (так называемый шеечный фактор бесплодия). По сообщению южноафриканского андролога Тинуса Крюгера для успешного зачатия в матку должно проникнуть не менее 10 млн сперматозоидов. Из матки сперматозоиды проникают в маточные, или фаллопиевы трубы. При выборе направления сперматозоиды движутся против тока жидкости. Ток жидкости в маточных трубах направлен от яичника в сторону матки, соответственно сперматозоиды движутся от матки в сторону яичника.
Непосредственно оплодотворение происходит в ампулярной части фаллопиевой трубы (расширяющаяся часть трубы, расположенная ближе к яичнику). Для осуществления слияния с яйцеклеткой сперматозоид должен преодолеть две оболочки: 1) лучистый венец (corona radiata), 2) блестящую оболочку (zona pellucida). Процесс преодоления оболочек яйцеклетки сперматозоидом и последующее слияние называют «пенетрацией». Для преодоления лучистого венца — слоя фолликулярных клеток, окружающих яйцеклетку, сперматозоид использует фермент гиалуронидазу, расположенный на поверхности головки и расщепляющий внеклеточный матрикс, соединяющий клетки лучистого венца. Один сперматозоид не способен разрушить лучистый венец, требуется воздействие большого количества сперматозоидов для разрыхления и рассеивания клеток лучистого венца. Сперматозоид, который первым доберется до блестящей оболочки, вероятнее всего осуществит оплодотворение.
Для преодоления блестящей оболочки zona pellucida сперматозоид использует ферменты акросомы. Акросома представляет собой видоизмененную лизосому — мембранный пузырек, заполненный различными литическими ферментами. Акросома располагается на переднем конце головки сперматозоида. После достижения сперматозоидом блестящей оболочки, рецепторы на головке сперматозоида взаимодействуют с лигандами на блестящей оболочке. После такого взаимодействия (узнавания) акросома сливается с внешней мембраной сперматозоида и её содержимое оказывается снаружи. Ферменты акросомы локально разрушают блестящую оболочку, что в совокупности с движением сперматозоида позволяет ему проникнуть под блестящую оболочку и оказаться в непосредственной близости к оолемме — цитоплазматической мембране яйцеклетки.
Существуют специфические рецепторы на поверхности сперматозоида и яйцеклетки, которые служат для осуществления слияния половых клеток. После слияния ядро сперматозоида, центриоль и его митохондрия оказываются внутри яйцеклетки. В течение короткого времени в яйцеклетке развивается так называемая кортикальная реакция, служащая для обеспечения блока полиспермии. Суть кортикальной реакции состоит в высвобождении содержимого кортикальных гранул, или везикул, в наружную от яйцеклетки среду. Кортикальные гранулы располагаются под поверхностью наружной мембраны яйцеклетки и содержат различные литические ферменты. После проникновения сперматозоида кортикальные везикулы сливаются с оолеммой (наружной мембраной яйцеклетки) и вышедшие из яйцеклетки ферменты модифицируют блестящую оболочку таким образом, что она становится недоступной для проникновения других сперматозоидов. Таким образом, после проникновения первого сперматозоида, остальные сперматозоиды не способны оплодотворить яйцеклетку. Механизм, препятствующий проникновению более одного сперматозоида в яйцеклетку, получил название «блок полиспермии». У млекопитающих блок полиспермии развивается в течение нескольких минут.
При проникновении двух сперматозоидов в яйцеклетку (что случается сравнительно редко) образуется триплоидный эмбрион, при дальнейшем развитии которого происходят различные нарушения в расхождении хромосом. Такие эмбрионы не являются жизнеспособными и, как правило, погибают в течение нескольких дней развития, однако в редких случаях могут имплантироваться в матку и дать начало беременности, которая обречена на прерывание. Вопреки распространенному заблуждению проникновение двух сперматозоидов в яйцеклетку не является причиной возникновения однояйцевых близнецов.
После проникновения сперматозоида в яйцеклетку образуется зигота — оплодотворенная яйцеклетка, одноклеточная стадия развития эмбриона. У человека стадия зиготы продолжается первые 26-30 часов развития. Зигота приступает к формированию ядер. Сперматозоид проникает в яйцеклетку на стадии второго деления мейоза (после овуляции мейоз в яйцеклетке останавливается на стадии метафазы II и возобновляется после проникновения сперматозоида). Зигота завершает мейоз и как следствие выделяет 2-е полярное тельце и формирует женский пронуклеус (женское ядро). Параллельно из материала ядра сперматозоида зигота формирует мужской пронуклеус (мужское ядро). Каждый из пронуклеусов зиготы имеет одинарный (гаплоидый) набор хромосом, в мужском пронуклеусе располагаются отцовские хромосомы, в женском пронуклеусе — материнские хромосомы. В микроскоп пронуклеусы в зиготе становятся видны спустя 13-15 часов после проникновения сперматозоида и исчезают спустя 20-21 час после проникновения сперматозоида. Сформировавшись на разных концах зиготы, пронуклеусы движутся навстречу друг другу (так называемое «сближение пронуклеусов»), после чего их оболочки растворяются и хромосомы выстраиваются в метафазную пластинку первого деления митоза. Таким образом, объединение отцовских и материнских хромосом происходит на стадии метафазы митоза зиготы. В отличие от зиготы иглокожих, в зиготе человека и других позвоночных животных не наблюдается слияния пронуклеусов с образованием единого ядра зиготы.
Дробление
В процессе оплодотворения яйцеклетка, а в последующем зигота, продолжает свое продвижение по маточной трубе в сторону матки. Этому способствуют сокращения мышечного слоя трубы и движения ресничек её эпителия. После образования зиготы начинается процесс её митотического деления, который носит название «дробление» (такое название деление зиготы получило потому, что общий размер эмбриона не увеличивается, и с каждым последующим делением дочерние клетки становятся все мельче). Размер эмбриона человека на стадиях зиготы и дробления одинаков и составляет около 130 мкм. Дробление у человека, как и всех млекопитающих, полное, равномерное и асинхронное. Асинхронное дробление означает, что дочерние клетки делятся не одновременно, в итоге эмбрион человека может содержать различное количество клеток (а не только 2 в степени n, как это характерно для большинства животных). Клетки эмбриона на стадии дробления называются бластомеры.
Период дробления продолжается около 3 дней. Первоначально все бластомеры эмбриона человека одинаковы, как по внешнему виду, так и по своей детерминации. Бластомеры не взаимодействуют друг с другом и удерживаются вместе лишь благодаря блестящей оболочке. Если блестящая оболочка по какой-то причине будет повреждена, то эмбрион рассыпется на отдельные группы клеток или индивидуальные клетки. В редких случаях это может приводить к формированию двух и более независимых эмбрионов, идентичных генетически. Такие эмбрионы дадут начало однояйцевым дихориальным близнецам (около одной трети случаев рождения всех однояйцевых близнецов).
К 4 дню развития, когда эмбрион состоит приблизительно из 12-16 клеток, бластомеры приобретают дифференциацию и образуют два клеточных слоя. Наружные бластомеры формирует так называемый трофобласт, а внутренние — чуть позже — эмбриобласт.
К 5 дню развития, дробящийся эмбрион формирует бластоцисту — стадию развития, характерную только для плацентарных млекопитающих. Бластоциста состоит из приблизительно 30 клеток в начале развития и приблизительно 200 клеток в конце развития. Бластоциста представляет собой полый шар размером 130-200 мкм, сформированный клетками трофобласта, внутри шара располагается группа клеток эмбриобласта, прикрепленная к одной из стенок.
Изредка бластоциста может нести два эмбриобласта, такой эмбрион даст начало однояйцевой двойне — однояйцевым монохориальным близнецам (около двух третей случаев рождения всех однояйцевых близнецов).
Имплантация
Для лучшего контакта с эндометрием бластоциста освобождается от блестящей оболочки (так называемый хэтчинг). После этого клетки трофобласта, поверхностного слоя бластоцисты, выбрасывают пальцевидные отростки для погружения в эндометрий, железы которого богаты питательным секретом. В то же время эндометрий продолжает утолщаться под влиянием прогестерона и в итоге окружает бластоцисту со всех сторон. Процесс имплантации происходит при тесном химическом и физическом взаимодействии бластоцисты и эндометрия. Хорионический гонадотропин, выделяемый клетками трофобласта, влияет также на жёлтое тело яичника, стимулируя выработку им прогестерона и препятствуя наступлению менструации.
Нарушения зачатия
В редких случаях нарушение кортикальной реакции приводит к оплодотворению яйцеклетки более чем одним сперматозоидом. Это называется полиспермией и обычно обусловливает нежизнеспособную зиготу. Закрытие просвета маточной трубы вследствие некоторых заболеваний ведет к препятствию для встречи сперматозоидов с яйцеклеткой и возникновению бесплодия. Нарушение транспорта зиготы в матку может приводить к имплантации бластоцисты в маточной трубе и развитию внематочной беременности. Нарушение реакций взаимодействия бластоцисты с эндометрием в процессе имплантации может обусловить ранний выкидыш ещё до того, как можно установить факт беременности.
См. также
Оплодотворение. Появление близнецов — урок. Биология, Человек (8 класс).
Оплодотворение
Во время полового акта сперматозоиды (около \(200\) миллионов) попадают во влагалище и передвигаются по направлению к матке. За несколько часов сперматозоиды достигают маточной трубы, где они могут встретить яйцеклетку. В маточной трубе происходит процесс оплодотворения.
Оплодотворение — процесс, в результате которого сперматозоид сближается с яйцеклеткой, и их ядра (в каждом из которых находится по \(23\) хромосомы), сливаются в единое целое — зиготу — клетку, ядро которой содержит \(46\) хромосом (по \(23\) от матери и отца) и которая, многократно делясь, даёт начало новому организму.
Пол будущего человека определяется в момент оплодотворения и зависит от того, какой сперматозоид оплодотворит яйцеклетку. В клетках имеются носители информации — хромосомы. Все яйцеклетки содержат хромосому Х. Половина сперматозоидов содержит хромосому Х, а другая половина содержит хромосому Y. Если яйцеклетку оплодотворит Х-содержащий сперматозоид, появится девочка; если Y-содержащий — родится мальчик.

Оплодотворение возможно в течение \(12\)–\(24\) часов после овуляции (так как в течение этого периода яйцеклетка сохраняет свою жизнеспособность). Способность к оплодотворению у сперматозоидов сохраняется от \(2\) до \(4\) суток.
Сам процесс оплодотворения происходит следующим образом. Множество сперматозоидов окружают яйцеклетку, и поверхность их головок вступает в контакт с её оболочками. При этом сперматозоиды выделяют фермент, увеличивающий проницаемость оболочек яйцеклетки. Обычно в яйцеклетку проникает и сливается с ней только один сперматозоид. После этого вокруг яйцеклетки образуется особая оболочка, препятствующая проникновению в неё ядер других сперматозоидов.

При овуляции и оплодотворении двух (или более) яйцеклеток образуются два или более плода. Это будущие разнояйцевые близнецы. Они не очень похожи друг на друга и даже могут быть разного пола.

Если два плода развиваются из одной яйцеклетки, то это однояйцевые близнецы. Они всегда одного пола и очень похожи друг на друга.


Источники:
Любимова З. В., Маринова К. В. Биология. Человек и его здоровье. 8 класс. — М.: Владос.
Лернер Г. И. Биология: Полный справочник для подготовки к ЕГЭ: АСТ, Астрель.
http://prelestnyi.ru/beremennost/1trimestr/4nedel-beremennosti/
http://www.beremka.ru/advice-for-pregnant/gemini/
Общая характеристика процесса оплодотворения.
⇐ ПредыдущаяСтр 3 из 8Следующая ⇒Оплодотворением называют слияние гаплоидного сперматозоида с гаплоидной яйцеклеткой, завершающееся объединением их ядер в единое диплоидное ядро оплодотворенного яйца – зиготы. В процессе оплодотворения сперматозоид выполняет две функции. Первая – активация яйца, побуждение его к началу развития. Эта функция не специфична для сперматозоида: в качестве активирующего фактора он может быть заменен рядом физических или механических агентов, способных спровоцировать развитие зародыша. Развитие яйцеклетки без участия сперматозоида называется партеногенезом. Другая функция сперматозоида, в выполнении которой он уже незаменим, – внесение в яйцеклетку отцовского генетического материала.
Взаимодействие половых клеток (гамет) в процессе оплодотворения можно разделить на три фазы: 1) дистантное взаимодействие, осуществляющееся на некотором расстоянии, до соприкосновения гамет; 2) контактное взаимодействие, происходящее при непосредственном соприкосновении поверхностей гамет; 3) процессы, протекающие после вхождения сперматозоида в яйцо (рис. 2.1).

Рис. 2.1. Процесс оплодотворения.
А – фаза дистантного взаимодействия; Б, В, Г – фаза контактного взаимодействия;
Д, Е, Ж, З – фаза синкариона. 1 – мембрана яйца; 2 – студенистая оболочка; 3 – бугорок оплодотворения; 4 — оболочка оплодотворения; 5 – центриоль.
1.1. Дистантное взаимодействие гамет направлено на повышение вероятности встречи сперматозоидов с яйцеклеткой. По большей части эти взаимодействия осуществляются через посредство хемотаксиса – движения сперматозоидов по градиенту концентрации некоторых веществ, выделяемых яйцеклеткой. Наличие хемотаксиса достоверно установлено для многих групп животных, особенно беспозвоночных: книдарий, моллюсков, иглокожих и полухордовых.
В движении сперматозоидов млекопитающих по верхним отделам яйцевода существенное значение имеет явление реотаксиса (способность двигаться против встречного тока жидкости в маточных трубах).
1.2. Контактное взаимодействие гамет начинает осуществляться с момента контакта сперматозоида с оболочками яйцеклетки (рис. 2.2). Первый этап этих взаимодействий получил название акросомной реакции. Иногда эта реакция может быть вызвана не только контактом с блестящей оболочкой яйцеклетки, но также соударением сперматозоида с любой твердой поверхностью или же при повышении концентрации Са2+. Внешнее, видимое при небольших увеличениях проявление этой реакции—выброс так называемой акросомной нити в сторону яйцевой оболочки. Тонкие электронно-микроскопические исследования сперматозоидов, фиксированных в период выбрасывания акросомной нити, показали следующее.
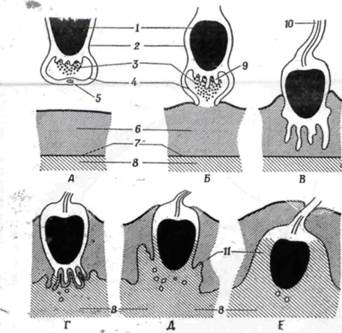
Рис. 2.2. Последовательные стадии соединения спермия с яйцом.
А. Б – раскрытие акросомного пузырька; В, Г – высвобождение лизирующих ферментов акросомы;
Д, Е – образование бугорка оплодотворения
Процесс начинается со слияния мембраны акросомы с наружной мембраной сперматозоида. Затем слившиеся мембраны разрываются, и происходит экзоцитоз содержимого акросомного пузырька. При этом из него изливаются спермолизины—ферменты, растворяющие оболочку яйцеклетки. После этого внутренний участок мембраны акросомы начинает быстро выпячиваться, в результате чего образуется одна или целый пучок так называемых акросомных трубочек (или микроворсинок), которые и выглядят при малом увеличении как нити. Акросомная микроворсинка растет в результате быстрой сборки фибриллярного сократительного белка актина, образующего ее структурную основу. Момент соприкосновения акросомной микроворсинки с блестящей оболочкой яйцеклетки — решающий для взаимного узнавания яйцeклeтки и сперматoзoидa.
Это узнавание осуществляется, в случае «правильной» встречи сперматозоида с яйцеклеткой того же вида, благодаря комплементарному взаимодействию особого белка (биндина), встроенного в мембрану акросомной микроворсинки (бывшая внутренняя мембрана акросомного пузырька) с соответственным рецептором на оболочке яйцеклетки. Даже у близких между собой видов биндины различаются по составу. Заключенные, таким образом, до акросомной реакции внутри акросомного пузырька биндины экспонируются (становятся доступными) для связывания рецепторами благодаря выворачиванию и росту акросомной микроворсинки.
Вслед за реакцией узнавания (образования комплекса между биндином и его рецептором в блестящей оболочке) оболочка яйцеклетки лизируется, после чего на ней образуется бугорок оплодотворения, направленный навстречу акросомной микроворсинке. Этот момент считается началом процесса активации яйцеклетки. Формирование бугорка оплодотворения, как и акросомной микроворсинки, сопровождается полимеризацией актина. Мембраны верхушки акросомной микроворсинки и бугорка оплодотворения сливаются между собой, и по образовавшемуся сквозному каналу содержимое сперматозоида (прежде всего ядро и по крайней мере одна из центриолей, но нередко также и хвостовая часть) переходит внутрь яйцеклетки. Участок мембраны сперматозоида встраивается в мембрану яйцеклетки и может сохраняться длительное время, иногда обнаруживаясь иммунологическими методами до стадии личинки (у морского ежа).
Быстрое повышение концентрации Са2+ также участвует в стимуляции синтеза белка и ДНК и обусловливает наиболее явный признак реакции активации яйцеклетки – экзоцитоз так называемых кортикальных альвеол (рис. 2.3). Это многочисленные пузырьки, содержащиеся в кортикальном (поверхностном) слое неоплодотворенной яйцеклетки. Со стимуляцией ионами Са2+ процессов экзоцитоза мы уже познакомились на примере экзоцитоза акросомного пузырька.
При экзоцитозе кортикальных альвеол из них высвобождаются в узкое пространство между плазматической мембраной яйцеклетки и плотно примыкающей к ней желточной оболочкой следующие вещества: 1) протеолитический фермент, разрывающий связи между плазматической мембраной и желточной оболочкой,— вителлиновая деламиназа; 2) протеолитический фермент, освобождающий осевшую на блестящей оболочке сперму от связей с этой оболочкой,— сперм-рецепторная гидролаза; 3) гликопротеид, втягивающий воду в пространство между желточной оболочкой и плазматической мембраной и вызывающий тем самым их расслоение: в результате между желточной оболочкой и плазматической мембраной яйцеклетки возникает обширное пространство, называемое перивителлиновым. Образование перивителлинового пространства — наиболее отчетливый признак активации яйцеклетки; 4) фактор, способствующий затвердеванию оболочки оплодотворения; 5) структурный белок гиалин, участвующий в формировании гиалинового слоя, располагающегося у многих яйцеклеток (например, морского ежа) над плазматической мембраной.
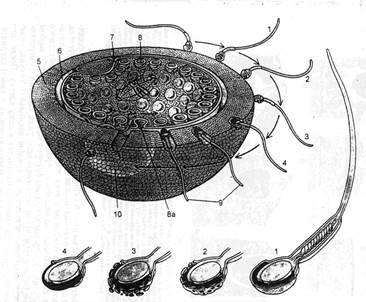
Рис. 2.3. Оплодотворение.
1, 2, 3 – стадии акросомной реакции; 5 – блестящая зона; 6 – перивителлиновое пространство;
7 – плазматическая мембрана; 8 – кортикальная гранула; 9 – вождение спермия в яйцеклетку;
10 – зонная реакция.
Одновременно происходит сборка и перераспределение элементов цитоскелета в кортикальном слое яйцеклетки. Кортикальный слой в результате приобретает сократимость, необходимую для осуществления делений дробления. Образование оболочки оплодотворения надежно предохраняет яйцеклетку от проникновения излишних сперматозоидов — полиспермии.
В первые секунды после контакта гамет резко повышается проницаемость плазматической мембраны яйцеклетки для внешнего Na+, что приводит к падению трансмембранного потенциала яйцеклетки от отрицательного (порядка —60 мВ) до слабо положительного (около +10 мВ). Это падение потенциала осуществляет так называемый быстрый блок полиспермии, так как в яйцеклетки с положительным трансмембранным потенциалом дополнительные сперматозоиды проникнуть уже не могут.
Таким образом, активация яйцеклетки — чрезвычайно быстрая и широкая по охвату реакция, вовлекающая в себя буквально все компоненты яйца.
1.3. Сперматозоид внутри яйца (фаза синкариона).
У большинства животных сперматозоид входит в яйцеклетку целиком, включая хвостовую часть; у некоторых видов жгутик остается на поверхности. Но, и оказавшись внутри яйцеклетки, жгутик сперматозоида не играет никакой роли в дальнейшем движении последнего. Сперматозоид сразу же поворачивается шейкой по ходу дальнейшего движения; вокруг центриоли возникает характерное «полярное сияние», образованное микротрубочками. Хроматин в ядре сперматозоида деспирализуется. Ядро сперматозоида называют теперь мужским пронуклеусом. Хроматин ядра яйцеклетки после завершения делений мейоза тоже деспирализуется. Это ядро называется женским пронуклеусом.
Прежде чем сблизиться, пронуклеусы проделывают сложные движения («танец пронуклеусов»). Сначала мужской пронуклеус движется внутрь яйца перпендикулярно поверхности и независимо от положения женского пронуклеуса. Этот отрезок пути называют «дорожкой проникновения». Затем оба пронуклеуса движутся навстречу друг другу по «дорожке копуляции». Движение мужского пронуклеуса осуществляется, по-видимому, благодаря «отталкиванию» растущих микротрубочек полярного сияния от поверхностного слоя яйцеклетки.
После сближения пронуклеусов наступает кариогамия — объединение их хромосомных наборов. Кариогамия происходит всегда только после завершения яйцеклеткой делений созревания (у большинства животных именно вхождение сперматозоида в яйцеклетку стимулирует завершение этих делений). У тех немногих видов, где сперматозоид проникает в уже зрелую яйцеклетку (например, у морского ежа), кариогамия выражается в непосредственном слиянии пронуклеусов; образуется единое ядро зиготы. В тех случаях, когда между вхождением сперматозоида и кариогамией проходит более длительный срок, оболочки пронуклеусов растворяются еще до их сближения, и хромосомы спирализуются. Тогда кариогамия выражается в том, что хромосомы обоих пронуклеусов располагаются в одной плоскости—плоскости метафазной пластинки 1-го митотического деления оплодотворенного яйца.
2. Ооплазматическая сегрегация – перемещение компонентов яйца после оплодотворения и формирование специфических участков («полей»), детерминирующим в дальнейшем развитие определенных частей зародыша.
Непосредственно после проникновения сперматозоида (или воздействия партеногенетического агента) начинаются интенсивные перемещения цитоплазмы яйцеклетки (ооплазмы). Иногда при этом происходит расслоение, отмешивание различных составных частей ооплазмы, что обозначается как ооплазматическая сегрегация. В ходе этого процесса намечаются основные, хотя и далеко не все, элементы пространственной организации зародыша.
3. Партеногенез.
Как уже говорилось, яйца многих животных могут быть активированы естественно или искусственно, без помощи сперматозоида. Развитие без участия сперматозоида называют партеногенезом. Естественный партеногенез типичен для летних поколений некоторых ракообразных и коловраток; он обнаружен у пчел, ос, ряда чешуекрылых, а из позвоночных — у некоторых видов ящериц и змей.
У млекопитающих также отмечались случаи вступления яйцеклеток на путь партеногенетического развития либо самопроизвольно, либо под влиянием различных активирующих агентов, например электростимуляции, теплового шока, этанола. Однако развитие таких зародышей всегда останавливалось на ранних стадиях развития. В некоторых случаях спонтанного партеногенеза дробящиеся зародыши становятся источниками опухолей яичника — тератом, в которых могут развиваться зачатки органов. Полноценное развитие партеногенетиков у млекопитающих невозможно потому, что в женских хромосомах заблокированы (в результате метилирования) определенные участки, содержащиеся в мужских хромосомах. Именно поэтому самец не может быть заменен у млекопитающих партеногенетическим агентом.
Лишь в редких случаях партеногенетически развивающиеся организмы являются гаплоидами (таковы самцы медоносной пчелы), В большинстве случаев после партеногенетической активации яйцеклетки в ней восстанавливается диплоидный набор хромосом.
Разновидностью партеногенеза является гиногенез — оплодотворение спермой другого (родственного) вида, которая лишь активирует яйцеклетку, но не вносит свой генетический материал в геном зародыша. Например, яйца серебряного карася могут быть стимулированы спермой сазана; плотвы, обыкновенного карася. В популяциях гиногенетических животных встречаются только самки. Имеются данные, что гиногенез может быть вызван искусственно термошоком для облучением яйцеклетки.
Андрогенез – явление, обратное партеногенезу, т.е. развитие яйцеклетки с участием только мужского ядра. Известны случаи естественного андрогенеза; андрогенетики встречаются у табака и кукурузы, иногда у тутового шелкопряда.
Андрогенез может быть вызван и искусственно. Еще в начале 19 века были поставлены опыты по оплодотворению фрагментов яиц морского ежа, лишенных собственного ядра. Такая разновидность искусственного андрогенеза, когда оплодотворяется фрагмент яйца, называется мерогонией.
II. Методические указания к выполнению лабораторной работы.
1. Изучить стадии оплодотворения по методическому пособию.
2. Ознакомиться с механизмами дистантного и контактного взаимодействия спермиев и яйцеклетки.
2.1. Зарисовать схему фазы дистантного взаимодействия (планшет № 2.1 «Дистантное и контактное взаимодействие спермиев и яйцеклетки»). Указать процесс капацитации сперматозоида (зарисовать рецепторы головки сперматозоида, процесс отделения углеводов с поверхности головки, процесс связывания рецепторов сперматозоида с НАГ-рецепторами).
2.2. Зарисовать фазу контактного взаимодействия. Отметить процесс связывания рецепторов сперматозоида с рецепторами яйцеклетки, процесс проникновения сперматозоида через оболочку яйцеклетки.
3. Ознакомиться с этапами акросомной реакции спермиев и кортикальной реакцией яйцеклетки по методическому пособию.
3.1. Зарисовать схему оплодотворения с указанием фазы контактного взаимодействия и синкариона (планшет № 2.2 «Оплодотворение; дробление»). Отметить период созревания, указать редукционные тельца. Рассмотреть и описать формирование мужского и женского пронуклеуса. Выделить оболочку оплодотворения.
4. Изучить процесс синкариона по методическому пособию.
4.1. Изучить под микроскопом и зарисовать препарат № 5.
Препарат № 5. Оплодотворение яйцеклетки. Яйцо аскариды с внедрившимся сперматозоидом (рис. 2.4).

Рис.2.4. Оплодотворение яйца аскариды:
1 – головка проникшего в яйцеклетку сперматозоида.
Препарат представляет собой группу яйцеклеток аскариды. Выберем при малом увеличении и поставим в центр поля зрения клетки, в которых отчетливо различимо содержимое. Переменив малое увеличение на большое, рассмотрим в них мелкозернистую цитоплазму и два ядра: одно более рыхлое, нередко в состоянии митоза – это женское ядро (яйцеклетка), другое более компактное, нередко еще сохраняющее треугольную форму, – это еще не вполне разбухшая головка спермия – мужское ядро. Эти ядра носят название пронуклеусов. Следовательно, здесь зафиксирован момент непосредственно после внедрения спермия в яйцо. В отдельных яйцеклетках между наружным краем протоплазмы и оболочкой сохранилось еще мелкое образование – направительное тельце.
Зарисуйте несколько клеток при большом увеличении.
4.2. Изучить под микроскопом и зарисовать препарат № 6.
Препарат № 6. Синкарион.Матка аскариды с оплодотворенными яйцеклетками (рис.2.5).
Препарат представляет собой поперечный разрез матки аскариды, набитой яйцеклетками. Последние окружены толстыми оболочками. Некоторые яйцеклетки еще не оплодотворены, в другие уже проникли сперматозоиды.
Изучаемый препарат фиксирует дальнейший этап оплодотворения: сближение и соединение женского и мужского ядра.
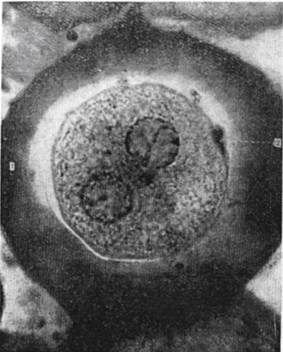
Рис. 2.5. Образование синкарионав в яйце аскариды:
1 – оболочка яйцеклетки; 2 – второе направительное тельце.
При малом увеличении, а еще отчетливее при большом, мы различаем в отдельных клетках соприкасающиеся, но еще лежащие отдельно ядра, в других оболочки ядер уже растворились и хромосомы объединились в общую группу.
Зарисуйте 2–3 наиболее типичные клетки при большом увеличении.
5. Ознакомиться с механизмом партеногенеза по методическому пособию.
6. Сдать отчет преподавателю и защитить его.
III. Содержание отчета.
Отчет должен быть представлен на отдельных листах формата А4 или в альбоме.
Отчет должен содержать:
1. Цель работы.
2. Краткое описание фаз оплодотворения.
3. Результаты исследований (микроскопическое изучение препаратов) и их анализ (с указанием использованных микроскопов, их увеличения, других приборов и материалов).
4. Результаты выполнения индивидуального задания (определение и описание «слепого» препарата).
5. Выводы.
Отчет на листе формата А4 сдается в конце работы преподавателю.
IV. Контрольные вопросы.
1. Перечислите стадии оплодотворения.
2. В чем заключается молекулярный механизм акросомной реакции?
3. В чем заключается механизм кортикальной реакции?
4. Что такое ооплазматическая сегрегация.
5. Объясните биологическую сущность партеногенеза.
Рекомендуемая литература.
1. А.В.Белоусов. Биология индивидуального развития., 1983.
2. К.Г.Газарян. Биология индивидуального развития животных.,1983.
3. О.В.Волкова. Атлас. Гистология, цитология, эмбриология, 1996.
4. С.Л.Кузнецов. Атлас эмбриологии, 2002.
ЛАБОРАТОРНОЕ ЗАНЯТИЕ № 3.
21. Оплодотворение. Биология. Общая биология. 10 класс. Базовый уровень
21. Оплодотворение
Вспомните!
Какой набор хромосом имеет зигота?
Для каких животных характерно наружное оплодотворение?
У каких организмов существует двойное оплодотворение?
Для осуществления полового размножения организму недостаточно просто сформировать половые клетки – гаметы, надо обеспечить возможность их встречи. Процесс слияния сперматозоида и яйцеклетки, сопровождающийся объединением их генетического материала, называют оплодотворением. В результате оплодотворения образуется диплоидная клетка – зигота, активация и дальнейшее развитие которой приводит к формированию нового организма. При слиянии половых клеток разных особей осуществляется перекрёстное оплодотворение, а при объединении гамет, продуцируемых одним организмом, – самооплодотворение.
Существует два основных типа оплодотворения – наружное (внешнее) и внутреннее.
Наружное оплодотворение. При наружном оплодотворении половые клетки сливаются вне организма самки. Например, рыбы мечут икру (яйцеклетки) и молоку (сперму) прямо в воду, где происходит наружное оплодотворение. Подобным образом осуществляется размножение у земноводных, многих моллюсков и некоторых червей. При наружном оплодотворении встреча яйцеклетки и сперматозоида зависит от самых разных факторов внешней среды, поэтому при таком типе оплодотворения организмы обычно образуют огромное количество половых клеток. Например, озёрная лягушка откладывает до 11 тыс. яиц, атлантическая сельдь вымётывает около 200 тыс. икринок, а рыба-луна – почти 30 млн.
Внутреннее оплодотворение. При внутреннем оплодотворении встреча гамет и их слияние происходит в половых путях самки. Благодаря согласованному поведению самца и самки и наличию специальных совокупительных органов мужские половые клетки поступают непосредственно в женский организм. Так происходит оплодотворение у всех наземных и некоторых водных животных. В этом случае вероятность успешного оплодотворения высока, поэтому половых клеток у таких особей гораздо меньше.
Количество половых клеток, которые образует организм, зависит также от степени заботы родителей о потомстве. Например, треска вымётывает 10 млн икринок и никогда не возвращается к месту кладки, африканская рыбка тиляпия, вынашивающая икру во рту, – не более 100 икринок, а млекопитающие, обладающие сложным родительским поведением, обеспечивающим заботу о потомстве, рождают всего одного или нескольких детёнышей.
У человека, как и у всех остальных млекопитающих, оплодотворение происходит в яйцеводах, по которым яйцеклетка движется по направлению к матке. Сперматозоиды преодолевают огромное расстояние до встречи с яйцеклеткой, и лишь один из них проникает в яйцеклетку. После проникновения сперматозоида яйцеклетка формирует на поверхности толстую оболочку, непроницаемую для остальных сперматозоидов.
Если оплодотворение произошло, яйцеклетка завершает своё мейотическое деление (§ 20) и два гаплоидных ядра сливаются в зиготе, объединяя генетический материал отцовского и материнского организмов. Образуется уникальная комбинация генетического материала нового организма.
Яйцеклетки большинства млекопитающих сохраняют способность к оплодотворению в течение ограниченного времени после овуляции, как правило, не более 24 часов. Сперматозоиды, покинувшие мужскую половую систему, живут тоже очень недолго. Так, у большинства рыб сперматозоиды погибают в воде уже спустя 1–2 минуты, в половых путях кролика живут до 30 часов, у лошадей 5–6 суток, а у птиц до 3 недель. Сперматозоиды человека во влагалище женщины гибнут спустя 2,5 часа, но те, которые успевают добраться до матки, сохраняют жизнеспособность в течение двух и более суток. Существуют в природе и исключительные случаи, например сперматозоиды пчёл сохраняют способность к оплодотворению в семяприёмнике самок в течение нескольких лет.
Оплодотворённая яйцеклетка может развиваться в теле материнского организма, как это происходит у плацентарных млекопитающих, или во внешней среде, как у птиц и пресмыкающихся. Во втором случае она покрывается специальными защитными оболочками (яйца птиц и пресмыкающихся).
У некоторых видов организмов встречается особая форма полового размножения – без оплодотворения. Такое развитие называют партеногенезом (от греч. partenos – девственница, genesis – возникновение) или девственным развитием. В этом случае дочерний организм развивается из неоплодотворённой яйцеклетки на основе генетического материала одного из родителей, и образуются особи только одного пола. Естественный партеногенез даёт возможность резкого увеличения численности потомства и существует в тех популяциях, где контакт разнополых особей затруднён. Партеногенез встречается у животных разных систематических групп: у пчёл, тлей, низших ракообразных, скальных ящериц и даже у некоторых птиц (индеек).
Одним из главных механизмов, который обеспечивает оплодотворение строго внутри вида, является соответствие числа и строения хромосом женских и мужских гамет, а также химическое сродство цитоплазмы яйцеклетки и ядра сперматозоида. Даже если чужеродные половые клетки и соединяются при оплодотворении, это, как правило, приводит к ненормальному развитию зародыша или к рождению стерильных гибридов, т. е. особей, неспособных к деторождению.
Двойное оплодотворение. Особый тип оплодотворения характерен для цветковых растений. Он был открыт в конце XIX в. русским учёным Сергеем Гавриловичем Навашиным и получил название двойного оплодотворения (рис. 67).
Во время опыления пыльца попадает на рыльце пестика. Пыльцевое зерно (мужской гаметофит) состоит всего из двух клеток. Генеративная клетка делится, образуя два неподвижных спермия, а вегетативная клетка, прорастая внутрь пестика, формирует пыльцевую трубку. В завязи пестика развивается женский гаметофит – зародышевый мешок с восемью гаплоидными ядрами. Два из них сливаются, формируя центральное диплоидное ядро. В результате дальнейшего деления цитоплазмы зародышевого мешка образуется семь клеток: яйцеклетка, центральная диплоидная клетка и пять вспомогательных.
Рис. 67. Двойное оплодотворение у цветковых растений
После того как пыльцевая трубка прорастает в основание пестика, спермии, находящиеся внутри неё, проникают в зародышевый мешок. Один спермий оплодотворяет яйцеклетку, – возникает диплоидная зигота; из неё в дальнейшем развивается зародыш. Другой спермий сливается с ядром крупной центральной диплоидной клетки, образуя клетку с тройным хромосомным набором (триплоидную), из которой затем формируется эндосперм – питательная ткань для зародыша. Таким образом, у покрытосеменных растений в оплодотворении участвует два спермия, т. е. осуществляется двойное оплодотворение.
Искусственное оплодотворение. Большое значение в современном сельском хозяйстве имеет искусственное оплодотворение, приём, который широко применяется в селекции при выведении и улучшении пород животных и сортов растений. В животноводстве при помощи искусственного осеменения можно получить многочисленное потомство от одного выдающегося производителя. Сперма таких животных хранится в специальных низкотемпературных условиях и сохраняет жизнеспособность в течение долгого времени (десятки лет).
Искусственное опыление в растениеводстве позволяет осуществлять определённое, заранее запланированное скрещивание и получать сорта растений с необходимым сочетанием родительских свойств.
В современной медицине при лечении бесплодия используется искусственное оплодотворение спермой донора и экстракорпоральное (внетелесное) оплодотворение – метод, разработанный впервые в 1978 г. и известный под названием «ребёнок из пробирки». Этот метод заключается в оплодотворении яйцеклеток вне организма и последующем переносе их назад в матку для продолжения нормального развития.
К 2010 г. с помощью экстракорпорального оплодотворения было зачато уже около 4 млн детей. Однако использование донорской спермы, донорских яйцеклеток и даже суррогатных матерей порождает целый ряд этических и социальных проблем. Многие люди, опираясь на религиозные и моральные соображения, выступают против любых вмешательств в размножение человека, в том числе против экстракорпорального и искусственного оплодотворения.
Вопросы для повторения и задания
1. Что такое оплодотворение?
2. Какие типы оплодотворения вы знаете?
3. В чём заключается процесс двойного оплодотворения?
4. Каково значение искусственного оплодотворения в растениеводстве и животноводстве?
Подумайте! Выполните!
1. Как вы считаете, в чём преимущество двойного оплодотворения у покрытосеменных растений по сравнению с оплодотворением у голосеменных?
2. Достаточно ли знать, что в размножении участвует только одна особь, чтобы сделать вывод о том, что это размножение – бесполое?
3. Объясните, почему при экстракорпоральном оплодотворении часто рождаются близнецы.
4. Организуйте и проведите дискуссию «Экстракорпоральное оплодотворение: за и против».
Работа с компьютером
Обратитесь к электронному приложению. Изучите материал и выполните задания.
Повторите и вспомните!
Растения
Опыление. Двойному оплодотворению у цветковых растений предшествует опыление – перенос пыльцы (пыльцевых зёрен) на рыльце пестика. Опыление осуществляется различными способами. Если пыльца цветка попадает на рыльце пестика этого же цветка, происходит самоопыление. Перенос пыльцы на рыльце пестика другого цветка называют перекрёстным опылением.
Самоопыление характерно для небольшого числа цветковых растений. Учёные считают, что самоопыление возникло вторично, когда какие-то обстоятельства стали препятствовать осуществлению перекрёстного опыления. Биологически самоопыление менее выгодно, поскольку при этом не происходит обмен генетической информации между различными особями вида.
Перекрёстное опыление распространено у покрытосеменных растений гораздо шире, чем самоопыление. Биологически перекрёстное опыление более благоприятно, чем самоопыление, потому что оно даёт возможность объединять генетическую информацию разных особей. Появляются потомки, отличающиеся от родительских особей. Это способствует приспособлению вида к изменяющимся условиям обитания.
Перекрёстное опыление может осуществляться различными способами. Условно их можно разделить на две группы: абиотическое опыление (при помощи ветра или воды) и биотическое (при помощи животных). В роли опылителей могут выступать разные животные: насекомые, птицы, млекопитающие.
Данный текст является ознакомительным фрагментом.Читать книгу целиком
Поделитесь на страничкеСледующая глава >
1. Оплодотворение и развитие плодного яйца. Акушерство и гинекология: конспект лекций
1. Оплодотворение и развитие плодного яйца
Оплодотворение – процесс соединения мужской и женской половых клеток. Оно происходит в ампулярной части маточной трубы. С этого момента начинается беременность.
Миграция оплодотворенного яйца
Оплодотворенное дробящееся яйцо продвигается по трубе в сторону матки и на 6–8-ой день достигает ее полости. Продвижению яйца способствуют перистальтические сокращения маточных труб, а также мерцание ресничек эпителия.
Имплантация оплодотворенного яйца
Слизистая оболочка матки ко времени попадания в полость матки оплодотворенного яйца резко утолщенная и рыхлая. В эндометрии вследствие влияния гормона желтого тела накапливается гликоген. Слизистая оболочка матки во время беременности называется децидуальной, или отпадающей оболочкой. Оплодотворенное яйцо, наружный слой которого представляет собой трофобласт, благодаря наличию протеолитических ферментов расплавляет децидуальную оболочку, погружается в ее толщу и прививается.
Плацента
В конце 1-го месяца беременности плодное яйцо со всех сторон окружено ворсинами хориона, которые вначале не имеют сосудов. Постепенно происходит васкуляризация хориона: в его ворсины врастают сосуды зародыша. На 2–3-м месяце беременности начинается атрофия ворсин хориона на одном полюсе плодного яйца, обращенном в полость матки. На противоположном участке хориона, погруженном в слизистую оболочку, ворсины пышно разрастаются и в начале 4-го месяца превращаются в плаценту. Помимо ворсин хориона, составляющих основную массу плаценты, в ее формировании принимает участие децидуальная оболочка матки (материнская часть плаценты). Плацента выделяет в материнский организм сложный комплекс гормонов и биологически активных веществ. Особое значение имеет прогестерон, способствующий развитию и сохранению беременности. Для развития беременности большое значение имеют также эстрогенные гормоны: эстрадиол, эстриол и эстрон. К концу беременности плацента имеет диаметр 15–18 см, толщину 2–3 см и массу 500–600 г. В плаценте различают две поверхности: внутреннюю (плодовую) и наружную (материнскую). На плодовой поверхности, покрытой водной оболочкой, проходят сосуды, радиально расходящиеся от пуповины. Материнская поверхность состоит из 15–20 долек. Плацента осуществляет функцию обмена веществ между матерью и плодом, барьерную функцию, а также является мощной железой внутренней секреции. Материнская кровь изливается в межворсинчатое пространство и омывает ворсины хориона. Кровь матери и плода не смешивается.
Пуповина
Представляет собой шнуровидное образование, в котором проходят две артерии и одна вена. По артериям течет венозная кровь от плода к плаценте, по вене притекает к плоду артериальная кровь. Прикрепление пуповины может быть центральным, эксцентричным, краевым или оболочечным. Нормальная длина пуповины в среднем 50 см. Послед образуется из плаценты, пуповины, оболочек плода (амниона и хориона) и изгоняется из матки после рождения плода.
Околоплодные воды
Образуются в результате секреции эпителием амниона, транссудации из крови матери и деятельности почек плода. К концу беременности накапливается примерно 1–1,5 л вод. В водах содержатся гормоны, белок в количестве 2–4 г/л, ферменты, макро– и микроэлементы, углеводы и другие вещества.
Данный текст является ознакомительным фрагментом.Читать книгу целиком
Поделитесь на страничкеСледующая глава >
Оплодотворение [Половой процесс] — это, что такое, какие, определение, значение, доклад, реферат, конспект, сообщение, вики — WikiWhat
Этапы оплодотворения
Механизм оплодотворения яиц сперматозоидами — сложный процесс, включающий следующие этапы:
- Физический контакт гамет.
- Акросомная реакция со стороны сперматозоида, проникающего в яйцо.
- Кортикальная реакция со стороны яйца — цитоплазматический процесс активации яйца.
- Образование пронуклеусов: преобразование ядра спермия в мужской пронуклеус и формирование женского пронуклеуса.
- Сингамия (непосредственно оплодотворение): сближение мужского и женского пронуклеусов и слияние гаплоидных ядер. В результате образуется диплоидное ядро зиготы (синкариона) — зародышевый пузырёк.
- Начало дробления.
Акросомная реакция
При акросомной реакции (рис. 40) происходит разрушение передней мембраны акросомного пузырька, вытягивание его задней стенки с преобразованием в акросомную трубочку, которая контактирует с поверхностью яйца и внедряется в цитоплазму (на поверхности трубочки находится гиалуронидаза). Ядро и проксимальная центриоль переходят по трубочке в цитоплазму.
Кортикальная реакция
При соприкосновении акросомной трубочки с цитоплазмой яйца возникает кортикальная реакция (рис. 41) — выброс содержимого кортикальных вакуол, образование оболочки оплодотворения.
Верхние части рисунка 41 показывают отношение между кортикальными гранулами, клеточной мембраной и желточной оболочкой в яйце до (А) и после (Б) оплодотворения. После оплодотворения материал кортикальных гранул, по-видимому, соединяется с желточной оболочкой и образует вместе с ней оболочку оплодотворения.
Нижние части рисунка 41 иллюстрируют динамику кортикальной реакции и образования оболочки оплодотворения во времени (В — Е), а также характер распространения кортикальной реакции по поверхности яйца от места проникновения сперматозоида.
После кортикальной реакции области мужского и женского ядерного материала покрываются ядерной мембраной, формируя мужской и женский пронуклеус.
Образование пронуклеусов
Образование мужского и женского пронуклеусов на примере яйца аскариды изображено на рисунке 42. Видна оболочка оплодотворения яйцеклетки, под ней — полярное тельце.
Сингамия
Сингамия — это слияние мужского и женского пронуклеусов, при этом зародыш становится диплоидным. Зигота приобретает очень прочную оболочку оплодотворения.
Между оболочкой и цитоплазмой зиготы возникает перивителлиновое пространство (рис. 43), выполняющее следующие функции:
- блокирование полиспермии,
- пространство для развития зародыша,
- приобретение плавучести икринок рыб.
Электронно-микроскопические исследования показали, что многие стороны взаимодействия сперматозоида с яйцом у всех животных схожи, поскольку оно включает слияние мембран. Вокруг яйца и сперматозоида также образуется единая оболочка. После того как сперматозоид проник в яйцо и произошло слияние ядер, выделяется студенистая оболочка оплодотворения, препятствующая слиянию яйца с другими сперматозоидами. Непосредственным последствием оплодотворения наряду с прочими является мощная активация обмена яйцеклетки. При этом используются накопленные в желтке углеводы и синтезируются различные ферменты из мРНК и рибосом.
Существует три механизма оплодотворения у животных: наружное, наружно-внутреннее и внутреннее.
Наружное оплодотворение
У большинства водных беспозвоночных яйца и сперматозоиды выделяются в воду, где и происходит оплодотворение: сперматозоиды плывут к яйцам, привлекаемые, вероятно, особыми химическими веществами. При этом многие сперматозоиды погибают ещё до встречи с яйцами. Отсюда и высокая плодовитость животных, своего рода «страховой фонд», повышающий шансы на оплодотворение какой-то части яиц. Тем не менее, слияние гамет все же не совсем дело случая, поскольку сперматозоиды и яйца выделяются в одно время и в одном месте (массовый нерест). Это результат брачного поведения взрослых самцов и самок, причём серьёзную роль играют химическая (феромональная) стимуляция и синхронизация размножения такими периодическими явлениями, как суточные, лунные и приливно-отливные ритмы. Примером может служить процесс размножения морского многощетинкового червя палоло. Это тихоокеанский массовый вид, который до размножения живёт на дне, периодически поднимается наверх и роится в поверхностных слоях. Здесь самцы и самки выбрасывают в воду половые продукты — сперматозоиды и яйца, большая часть которых и сливается. Это явление строго периодично и совпадает с определённым временем года и фазами луны.
После наружного оплодотворения из яиц морских беспозвоночных часто развиваются подвижные планктонные личинки, живущие в толще воды и питающиеся фитопланктоном, обычно мелкими водорослями. Это облегчает выживание и расселение, уменьшает конкуренцию из-за пищи и предоставляет потомству разнообразие местообитаний. И все же процесс наружного оплодотворения очень расточителен, поскольку животным приходится производить огромное количество гамет, чтобы гарантировать развитие немногих зигот. Поэтому в процессе эволюции животных наружное оплодотворение сменяется наружно-внутренним или внутренним. Материал с сайта http://wikiwhat.ru
Наружно-внутреннее оплодотворение
Наружно-внутреннее оплодотворение состоит в том, что самец выводит сперму в виде капелек жидкости или в виде мешочка-сперматофора в наружную среду на субстрат, после чего его захватывает самка. Такое осеменение присуще наземным членистоногим.
Внутреннее оплодотворение
Внутреннее оплодотворение происходит во время копуляции, при которой сперма вводится самцом в половые пути самки либо с помощью специального копулятивного органа, либо опять же посредством сперматофора, который подвешивается к половому отверстию самки или же вводится в него. И в этом случае копуляции может предшествовать стимуляция самки определённым, характерным для каждого вида, «брачным» поведением самца. Иногда стимуляция необходима из-за агрессивного поведения самки, которая может даже съесть самца во время или после копуляции, как это бывает у пауков. Препятствием для межвидовой копуляции является форма копулятивного органа самца (у насекомых), соответствующая строению влагалища самки. Такое соответствие получило название принципа «ключа и замка». Это такой же видовой признак, как форма и окраска тела, число хромосом и т. д.
Картинки (фото, рисунки)
Рис. 40. Акросомная реакция у червя Saccoglossus kowalevskii
Рис. 41. Кортикальная реакция и образование оболочки оплодотворения. Описание рисунков А-Е смотрите в тексте статьи. 1 — эндоплазма; 2 — кортикальный слой; 3 — желточная оболочка; 4 — клеточная мембрана; 5 — оболочка оплодотворения; 6 — перивителлиновое пространство
Рис. 42. Образование мужского и женского пронуклеусов в яйце аскариды
Рис. 43. Перивителлиновое пространство между оболочкой и цитоплазмой в оплодотворённой яйцеклетке рыбы буффало
Почему у животных ведущих наземный образ жизни происходит внутреннее
Внутренние оополотаорение ю
Наружное оплодотворение кратко
У каких беспозвоночных внутреннее оплолдотворение
Основные этапы: оплодотаорение
В чём заключается биологическое значение процесса оплодотворения?
Какие существуют виды оплодотворения?
Какие фазы имеет процесс оплодотворения?
Каков механизм проникновения спермия в яйцо?
В чём проявляется реакция яйцеклетки на проникновение сперматозоида?
Какую роль выполняет перивителлиновое пространство?
Как взаимодействуют яйцо и сперматозоид после его проникновения в цитоплазму?
Что такое сингамия?