Гомологи, определение — Справочник химика 21
Разделение основных групп смолисто-асфальтовых веществ и количественное определение их проводятся на основании различной растворимости компонентов смолисто-асфальтовых веществ в разных растворителях и, в частности, на основании резко выраженного различия растворимости главных их составных частей (смол и асфальтенов) в низших гомологах метана. [c.463]Итак, данные разных методов не согласуются между собой преимущественным гомологом по данным рентгенографии является гомолог с и=34, а по данным хроматографии — гомолог с и=26. Напомним, что обычное завышение номера преимущественного гомолога, определенного методом рентгенографии, не выходит за пределы одного номера. В то же время по данным хроматографии количество гомолога с и=34 очень мало (0.84%). Если предположить, что кратные максимумы на рентгенограмме все-таки принадлежат разным фазам, то окажется, что одному из пиков соответствует гомолог /1=34, а другому пику — гомолог /1=17 (последний гомолог вообще отсутствует в составе смеси).

Антрагидрохинон и его гомологи, определение 7995 Антраниловая кислота, взаимодействие с Се 6273 Антрахинон анализ 6803. 7398 Антрахинон—диметил-анилин, цветная реакция на [c.350]
В табл. 3.6 приведены данные о некоторых параметрах дипольно-сегментального процесса этих гомологов, определенных в диапазоне от 50 Гц до 200 кГц при температурах на 25-30 °С выше их Гс. [c.31]
Каждому циклу нефтегазообразования свойственна своя нефтематеринская порода со специфическим для данного цикла составом ОВ. Эта специфика наследуется нефтью. Каждому циклу соответствует свой генотип нефти. Поэтому в основе прогнозирования качественного состава углеводородных флюидов должен лежать прежде всего генотип нефти, связанный с определенным циклом нефтегазообразования. В зависимости от специфики ОВ (гумусовой или сапропелевой его основы) и термобарических условий материнской породой будет генерироваться преимущественно газ или нефть.
Соединения включения с пустотами в виде каналов позволяют достаточно селективно разделять углеводороды, имеющие характерные для определенного гомологического ряда структурные элементы. Соединения включения с пустотами в виде ячеек иногда позволяют разделять два смежных гомолога (например, бензол и толуол) было предложено для этого также использовать соединения типа гидратов углеводородов. Соединения включения с пустотами в виде слоев чаще используют при избирательной сорбции в динамических системах, где наряду с молекулярными характеристиками компонентов заметную роль играет скорость переноса вещества в свободном объеме.
К аналогичным выводам пришли при изучении и высших гомологов.
 Правая ось позволяет судить о составе продуктов пиролиза, имеющих общую формулу С Н2 +2- Для этого из точки, соответствующей определенному времени контакта на кривой следует восстановить перпендикуляр до пересечения с вышележащей кривой. Проекция точки пересечения на правую ось соответствует количеству атомов углерода в молекуле углеводорода, полученного в результате пиролиза. [c.107]
Правая ось позволяет судить о составе продуктов пиролиза, имеющих общую формулу С Н2 +2- Для этого из точки, соответствующей определенному времени контакта на кривой следует восстановить перпендикуляр до пересечения с вышележащей кривой. Проекция точки пересечения на правую ось соответствует количеству атомов углерода в молекуле углеводорода, полученного в результате пиролиза. [c.107]Изучение кинетики окисления гомологов метана позволило открыть существование холодных пламен , двух точек воспламенения и, для определенной зоны температур (380—430 °С), некоторого отрицательного влияния температуры на скорость реакции (в этом интервале с повышением температуры уменьшается скорость окисления). Это, видимо, связано с тем, что образование ацетальдегида и свободного метильного радикала может привести (при указанных температурах) к цепным разветвленным реакциям, в то время как образование пропилена не сопровождается разветвлением цепи. [c.136]
Существует несколько способов рассмотрения количественного содержания индивидуальных углеводородов в нефтях. Наиболее простой из них — это определение абсолютной концентрации данного соединения в нефти. Однако при этом теряются весьма важные закономерности, связанные с концентрационным распределением изомеров. Кроме того, незначительные по своей абсолютной величине цифры (например, 0,02, 0,005% и т. д.), характеризующие концентрации отдельных углеводородов, плохо воспринимаются и плохо запоминаются. Более интересной является оценка относительного распределения изомеров или гомологов. Этот способ, предложенный в работе [8], весьма удобен для различных теоретических обобщений и дает наглядное представление о связи между строением алканов и их концентрацией в различных нефтях.
Наиболее простой из них — это определение абсолютной концентрации данного соединения в нефти. Однако при этом теряются весьма важные закономерности, связанные с концентрационным распределением изомеров. Кроме того, незначительные по своей абсолютной величине цифры (например, 0,02, 0,005% и т. д.), характеризующие концентрации отдельных углеводородов, плохо воспринимаются и плохо запоминаются. Более интересной является оценка относительного распределения изомеров или гомологов. Этот способ, предложенный в работе [8], весьма удобен для различных теоретических обобщений и дает наглядное представление о связи между строением алканов и их концентрацией в различных нефтях.
Работа 5. Определение бензола и его гомологов в их смеси. [c.210]
В случае церезинов также выявляются расхождения в номерах преимущественных гомологов, определенных с использованием методов рентгенографии и хроматографии. Вероятная причина расхождения все та же (см. раздел 6.2) она связана со стремлением молекул разной длины максимально плотно упаковаться в структуре твердого раствора. Следствием различных вариантов плотной упаковки молекул разной длины является образование сверхпериодических структур, разнообразие которых определяется гомологическим составом и характером распределения гомологов по числу и. Церезины Ц-67, Ц-80 и Ц-С могут бьггь охарактеризованы сверхпериодической ромбической ячейкой, состоящей из двух пакетов , каждый из которых включает три молекулярных слоя (см. рис. 65, б). [c.303]
раздел 6.2) она связана со стремлением молекул разной длины максимально плотно упаковаться в структуре твердого раствора. Следствием различных вариантов плотной упаковки молекул разной длины является образование сверхпериодических структур, разнообразие которых определяется гомологическим составом и характером распределения гомологов по числу и. Церезины Ц-67, Ц-80 и Ц-С могут бьггь охарактеризованы сверхпериодической ромбической ячейкой, состоящей из двух пакетов , каждый из которых включает три молекулярных слоя (см. рис. 65, б). [c.303]
Углеводороды. В технологии инертных газов и при их использовании в качестве защитных сред обычно требуется определять суммарное содержание углеводородов. Если среди углеводородов присутствуют метан и его гомологи, определение общей концентрации возможно практически только путем каталитического сожжения углеводородов в присутствии кислооода над платиной при 600° С или на нагретой окиси меди при 450°С. После сожжения углеводородов определяют количество образовавшейся углекислоты любым из описанных выше методов с последующим пересчетом результатов.
К ароматическим углеводородам, получаемым и перерабатываемым на нефтехимических заводах, относятся бензол, толуол и ксилол. Их получают каталитическим риформингом определенного нафтенового сырья. В меньшем масштабе при помощи специальных процессов получают и другие ароматические углеводороды — нафталин, его гомологи, а также ряд других конденсировапных ароматических углеводородов. [c.9]
Так как химический состав бензинов может определенным образом влиять на степень изомеризации гомологов иик-лонеитана в циклогексановые углеводороды, поэтому мы задались целью изучить влияние хлористого алюминия на степень изомеризации гомологов циклопентана, входящих в состав норийского бензина.
При изучении реакции алкилирования ацетиленом и его гомологами ароматических соединений, в частности фенолов , синтезированные дифенолы анализировали с помощью хроматографии в тонком слое окиси алюминия. Матовую стеклянную пластинку покрывали товарной хроматографической окисью алюминия в сухом виде (слой толщиной 0,5 мм, без применения фиксирующих средств). Дифенолы лучше всего разделялись элюэнтом, представляющим собой раствор этанола в бензоле в отношении 1 15. Хроматогргмму проявляли, используя пары иода. Для количественного определения компонентов был опробован метод измерения и сравнения площадей их пятен. Оказалось, что при хорошем разделении компонентов и при резких границах пятен этот метод расчета дает достаточно точные данные. Ошибка определения менее 6%. Этим методом были разделены дифенолы и их орто-пара-замещенные изомеры. Необходимо отметить, что в этой работе количество определяемого компонента было 10% и выше, поэтому о возможности применения метода для анализа микроколичеств судить трудно.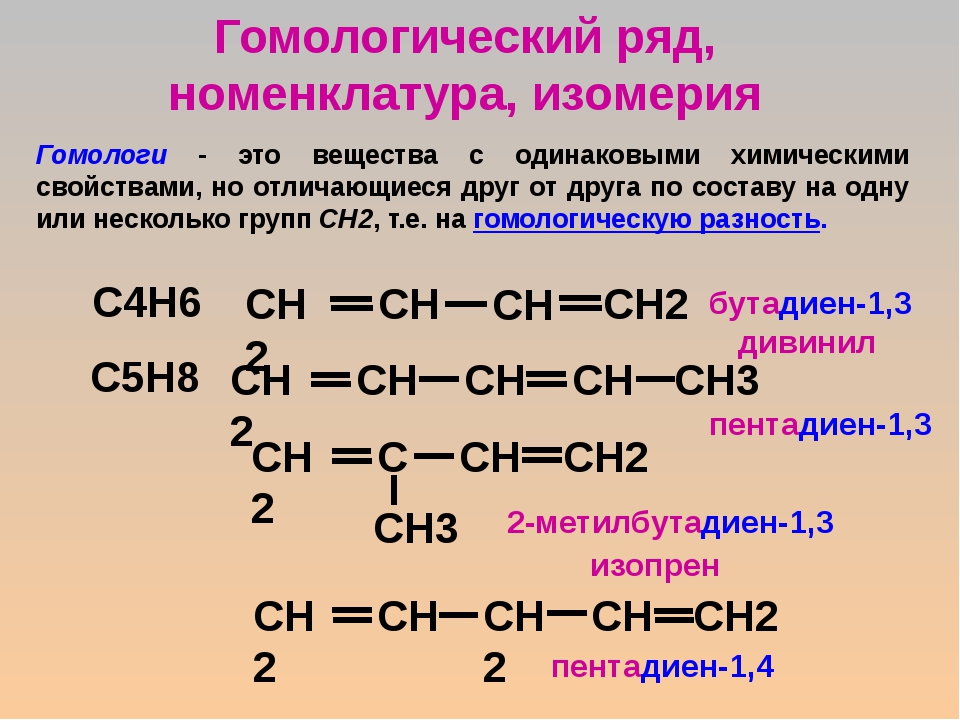
На количество образующегося кокса влияет не только химический состав основной массы сырья, но также присутствие небольших количеств асфальтовых соединений. Количество таких соединений может измеряться числом осмоления — количеством вещества, удаляемым серной кислотой [97], или коксовым числом (ASTM D 189-52). Последнее определение можно сделать более чувствительным, если находить коксовое число для 10 %-ной наиболее высококипящей фракции вещества. Если значение коксового числа превышает 0,12%, значит, нри крекинге будет образовываться избыточное количество кокса. В зависимости от характера сырья изменяют режим процесса, причем стараются добиться достаточно высокого выхода бензина при минимальном отложении кокса в аппаратуре. Выбор режима процесса следует связывать также с изменениями в стабильности фракций, которая зависит от соотношения между различными классами углеводородов и от соотношения между гомологами внутри определенного класса.
 Следует учесть, что, конечно, необходимые изменения в технологии зачастую незначительны. [c.309]
Следует учесть, что, конечно, необходимые изменения в технологии зачастую незначительны. [c.309] Определение серы в бензинах прямой гонки и в крэкинг-бензинаг состоит в определении активной серы, поскольку дисульфидная сера вполне нейтральна и не считается вредной, по крайней мере с точки зрения коррозии. Как известно, столь же безвредным считается тиофен и его гомологи. Так как обработка докторским раствором все жене может быть обставлена с количественным учетом результатов, н так как этот раствор делает бензин сладким , переводя меркаптаны в меркаптиды, а затем в дисульфиды, для определения меркаптанов необходимы иные методы анализа..
Относительно природы ароматических углеводородов керосина высказывались разные мнения. Данные пиролиза керосина и его ароматического экстракта сернистым газом (жидким) вполне определенно позволяют заключгггь о наличии почти исключительно лшого-замещенных гомологов и лишь в очепь небольшой степени однозамещенных с длинной боковой цепью (в пользу этого допущения говорят лишь косвенные доказательства). [c.204]
[c.204]
В случае советских нефтей почти не приходится иметь дела с определением ароматических углеводородов и метода для такой формы анализа пока что не существует, тш как старый способ Кре-мера — взбалтывание с равным объемом серной кислоты, приче ) наблюдается сокращеппе объема керосина — надо совсем оставить. Нет-никаких оснований полагать, что все ароматические углеводороды одинаково легко растворимы в креш ой серной кислоте. Наоборот, определенно известно, что некоторые из высших гомологов вовсе в пен нерастворимы, а многие, вроде одного из т1эиметилбензолов и этилтолуолов растворимы слабо. Применение же слабо дымящей кислоты, можпо сказать с уверенностью, откроет до 5% ароматических углеводородов там, где нет и 0,1%. [c.206]
Тяжелые углеводороды. Под этим термином понимаются прежде всего все непредельные углеводороды, затем бензол и его гомологи, отчасти пары жидких метановых углеводородов. Во всяг ом случае, только с некоторым приблия-гением можно понимать их как химически определенную группу. Для удаления их из газовой смеси пользуются связыванием их дымящей серной кислотой или бромной водой Первая растворяет непредельные углеводороды и ароматические. вторая связывает в ввде жидких бромидов, причем наблюдается постепенное обесцвечивание бромной воды (чтобы реактив не терял своей способности связывать непредельные соединения, в пипетке всегда должен быть свободный бром 2—4 г). На свету бром отчасти замещает водород в метановых углеводородах с другой сто-)оны, серная кислота растворяет отчасти и метановые углеводороды. Лоэтому при работе ио обоим методам, вообще говоря, не может быть строгого совпадения. Разница (несколько процентов) зависит от различных внешних, условий. Всегда надо указывать, какик реактивом произведено отделение тяжелых углеводородов. [c.384]
Для удаления их из газовой смеси пользуются связыванием их дымящей серной кислотой или бромной водой Первая растворяет непредельные углеводороды и ароматические. вторая связывает в ввде жидких бромидов, причем наблюдается постепенное обесцвечивание бромной воды (чтобы реактив не терял своей способности связывать непредельные соединения, в пипетке всегда должен быть свободный бром 2—4 г). На свету бром отчасти замещает водород в метановых углеводородах с другой сто-)оны, серная кислота растворяет отчасти и метановые углеводороды. Лоэтому при работе ио обоим методам, вообще говоря, не может быть строгого совпадения. Разница (несколько процентов) зависит от различных внешних, условий. Всегда надо указывать, какик реактивом произведено отделение тяжелых углеводородов. [c.384]
Вдвигая или выдвигал эту трубку, сообщение внутренности колбочки с наружной атмосферой можно выключить. После того как в тарированную колбочку влита навеска масла, ее снова взвешивают, определяя таким образом вес масла. Затем в нее вливают определенное количестю раствора пикриновой кислоты и выкачивают воздух при помощи насоса. Выдвинув трубку из пробки, разобщдют соединение-внутренности колбочки с воздухом и нагревают ее несколько часов на водяной бане при частом встряхивании. По охлаждении избыток ппкриновой кислоты оттитровывается щелочью или лучше едким барнтом в присутствии лакмоида в качестве индикатора. По количеству связанной пикриновой кислоты определяется количество нафталина. Так как и другие углеводороды тоже образуют пикраты . результат не может быть особенно точным. Напр., в нефтяной смоле содержатся метилнафталины, некоторые из которых жидки даже при 0° и друше гомологи его, обладающие близкими температурами кипения. Все эти гомологи тоже дают пикраты. Способ Кюстера таким образом не может претендовать на большую точность, если предварггтельно не выделить достаточно узкую фракцию нафталина, которую и надлежит обработать тем или иным способом. [c.424]
Затем в нее вливают определенное количестю раствора пикриновой кислоты и выкачивают воздух при помощи насоса. Выдвинув трубку из пробки, разобщдют соединение-внутренности колбочки с воздухом и нагревают ее несколько часов на водяной бане при частом встряхивании. По охлаждении избыток ппкриновой кислоты оттитровывается щелочью или лучше едким барнтом в присутствии лакмоида в качестве индикатора. По количеству связанной пикриновой кислоты определяется количество нафталина. Так как и другие углеводороды тоже образуют пикраты . результат не может быть особенно точным. Напр., в нефтяной смоле содержатся метилнафталины, некоторые из которых жидки даже при 0° и друше гомологи его, обладающие близкими температурами кипения. Все эти гомологи тоже дают пикраты. Способ Кюстера таким образом не может претендовать на большую точность, если предварггтельно не выделить достаточно узкую фракцию нафталина, которую и надлежит обработать тем или иным способом. [c.424]
Согласно Е. А. Рогозиной, С.Г. Неручеву, В.А. Успенскому (1974 г.), в верхней части осадочных пород генерация УВ в больших масштабах в результате термокаталитических процессов происходить не может (см. рис. 1), а значительные запасы газов в этой толще обусловлены перетоком их из нижней газовой зоны. Заметим при этом, что большинство исследователей стали считать необходимым детальное расчленение зоны катагенеза в соответствии с подразделениями процесса углефикации каменных углей. При этом следует указать, что одни и те же стадии катагенеза привязываются различными исследователями к разным глубинам (см. рис. 1), а отнесение слоев к той или иной стадии углефикации производится без всяких доказательств. Любопытно отметить, что представления о масштабах генерации УВГ и глубинах формирования газовых залежей у Е.А. Рогозиной, С.Г. Неручева и В.А. Успенского базируются на изучении конкретного разреза (рис. 5). Приведенные для этого разреза данные свидетельствуют об отсутствии какой-либо строгой закономерности в изменении содержания как метана, так и его газообразных гомологов каждый пласт характеризуется определенным содержанием указанных У В и подчас резко отличается от смежных пластов.
А. Рогозиной, С.Г. Неручеву, В.А. Успенскому (1974 г.), в верхней части осадочных пород генерация УВ в больших масштабах в результате термокаталитических процессов происходить не может (см. рис. 1), а значительные запасы газов в этой толще обусловлены перетоком их из нижней газовой зоны. Заметим при этом, что большинство исследователей стали считать необходимым детальное расчленение зоны катагенеза в соответствии с подразделениями процесса углефикации каменных углей. При этом следует указать, что одни и те же стадии катагенеза привязываются различными исследователями к разным глубинам (см. рис. 1), а отнесение слоев к той или иной стадии углефикации производится без всяких доказательств. Любопытно отметить, что представления о масштабах генерации УВГ и глубинах формирования газовых залежей у Е.А. Рогозиной, С.Г. Неручева и В.А. Успенского базируются на изучении конкретного разреза (рис. 5). Приведенные для этого разреза данные свидетельствуют об отсутствии какой-либо строгой закономерности в изменении содержания как метана, так и его газообразных гомологов каждый пласт характеризуется определенным содержанием указанных У В и подчас резко отличается от смежных пластов. Кривая изменения содержания рассматриваемых газов — это одна из возможных кривых указанных изменений. [c.7]
Кривая изменения содержания рассматриваемых газов — это одна из возможных кривых указанных изменений. [c.7]
Кратко остановимся на вопросе расчета состава газовой и жидкой фаз смесей углеводородов с надкритическими газовыми компонентами, такими как метан и его гомологи, яри высоких давлениях. Такие смеси в виде газоконденсатных и газонефтяных залегают на разных глубинах осадочной толщи земли. Из-за отсутствия теоретических методов расчета фазового равновесия таких смесей при высоких давлениях определение состава их равновесных фаз ведут по константам фазового равновесия углеводородов К ). Величина углеводорода I представляет собой отношение его мольных долей в равновесных газовой и жидкой фазах системы. Величина К зависит не только от температуры и давления системы и от природы углеводорода 1, но и от природы и концентрации всех других компонентов системы. Константы фазового равновесия углеводородов определяются по атласу констант, периодически публикуемому Американской ассоциацией для снабжения и переработки природного газа. Методы расчета состава фаз в углеводородных системах с помощью констант фазового равновесия подробно описаны в ряде работ [Е11ег1 С. К-, 1957 г. Степанова Г. С., 1974 и Намиот А. Ю., 1976 и др.]. [c.14]
Методы расчета состава фаз в углеводородных системах с помощью констант фазового равновесия подробно описаны в ряде работ [Е11ег1 С. К-, 1957 г. Степанова Г. С., 1974 и Намиот А. Ю., 1976 и др.]. [c.14]
В. А. Соколов, В. Ф. Симоненко и Н. Д. Гуляева (1977 г.), изучив газообразование у углей различных стадий углефикации, пришли к выводу, что интенсивность генерации газа углями последовательного углефикационного ряда протекает неравномерно, но в определенной последовательности, в которой различают три этапа первый этап (стадии углефикации Б—Д) отличается наиболее интенсивной генерацией газов, в составе которых доминирует (до 90%) двуокись углерода и присутствует метан. Второй этап (стадия Г—ОС) отличается образованием гомологов метана, содержание которых в отдельных случаях достигает 80%. На этом этапе реализуется значительная часть газопроизводящего и весь нефтепроизводящий потенциал гумусовых углей. Третий этап (стадии Г—А) характеризуется значительным выделением метана и прекращением образования его гомологов.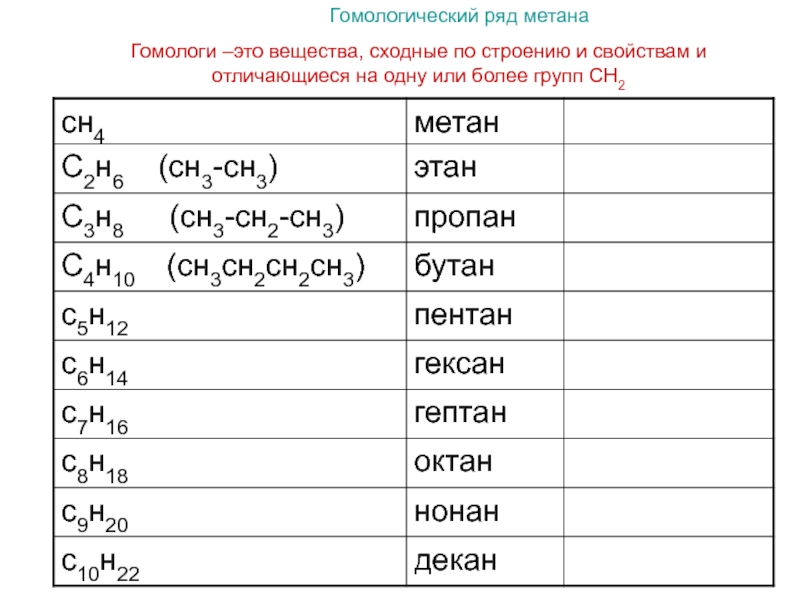 По мнению авторов, он соответствует главной [c.132]
По мнению авторов, он соответствует главной [c.132]
Химические методы разделения и исследования состава нефти основлваются на применении групповых реакций ее компонентов. В пределах даже широких фракций, таких как бензин ил1[ 1 еросан, по реакционной способности гомологи мало отличаются друг от друга, и поэтому химическими методами пх разделить трудно. С другой стороны, в любых фракциях различия между классами и группами соединений проявляются в заметной степени, в ряде случаев достаточной для аналитических целей. При обработке данного вещества определенным химическим реагентом в строго установленных условиях можно разделить смесь по типу молекул. Здесь, как и при исследовании физическими методами, наиболее надежные результаты получают прп работе с узкими фракциями. Когда предварительное разделение вещества на узкие фракции по техническим причинам невозможно, химической обработке должно все же предшествовать фракционирование, хотя бы и не очень четкое (па 30—50-градусные фракции). Тогда компоненты смеси, выделенные химическим методом, или компоненты, оставшиеся не затронутыми этой обработкой, исследуют в дальнейшем при помощи новой комбина-пии физических и химических методов. [c.87]
Тогда компоненты смеси, выделенные химическим методом, или компоненты, оставшиеся не затронутыми этой обработкой, исследуют в дальнейшем при помощи новой комбина-пии физических и химических методов. [c.87]
Эти углеводороды выделяют хроматографическим методом на силикагеле, поэтому в их составе содержатся и изопарафины, однако содержание последних невелико. Кроме того, нафтеновые углеводороды масляных фракций ефти являются смешанным.и, т. е. содержат в молекулах и па1рафиновые цепи. Нафтеновых углеводородов в негибридизираванном виде в высокомолекулярной части нефти, по имеющимся в литературе данным, вообще не со-де ржится. В работах Л. Г. Жердевой, Д. О. Гольд берг и других исследователей на основаиии определения элементного состава и физических свойств узких высококипящих фракций нафтеновых углеводородов показано, что в их составе наряду с гомологами циклогексана присутствуют и полициклические нафтены. Было установлено наличие в масляных фракциях бакинских нефтей нафтенов с 2, 3 и 4 циклами в молекуле. В работе Ф. Д. Россини показано, что число колец, содержащихся в молекулах нафтенов, зависит от пределов выкипания фракции. В легких масляных фракциях содержатся в основном гомологи циклогексана, в средних фракциях — алмилзамещенные нафтены с двумя и тремя циклами в молекуле, а в высококипящих фракциях обнаружены ди-, три- и тетрациклические конденсированные нафтеновые углеводороды. [c.9]
В работе Ф. Д. Россини показано, что число колец, содержащихся в молекулах нафтенов, зависит от пределов выкипания фракции. В легких масляных фракциях содержатся в основном гомологи циклогексана, в средних фракциях — алмилзамещенные нафтены с двумя и тремя циклами в молекуле, а в высококипящих фракциях обнаружены ди-, три- и тетрациклические конденсированные нафтеновые углеводороды. [c.9]
Для оценки результатов непрерывной ректификации, в частности смесей гомологов, большую пользу может оказать построение ступенчатой (идеализированной) диаграммы [242] (рис. 116). На ее основе можно построить столбиковую диаграмму фракционного состава смеси (рис. 117). Наглядные симметричные диаграммы Майер—Грольмана и Веселовского [243] также позволяют быстро проанализировать результаты разделения, полученные при разгонке по Энглеру или при аналитической ректификации многокомпонентных смесей, например моторных топлив, сланцевых масел, смол. По этому методу на ось у наносят значения температуры кипения, а на ось х (вправо и влево от оси у) — выход дистиллята в процентах от общего количества по фракциям, укладывающимся в температурные интервалы не уже 10 °С. На диаграмме получают площади (их обычно заштриховывают), напоминающие по форме репу или луковицу и дающие наглядное представление о результатах разгонки. Дополнительно слева на симметричной диаграмме приводят ряд чисел, соответствующих количествам дистиллята (в %), которые были получены с момента начала разгонки до определенной температуры справа на диаграмме наносят числа, показывающие выход дистиллята (в %) для определенных температурных интервалов. На диаграмме разгонки (рис. 118, 6 значение 180 С соответствует верхнему температурному пределу бензиновой фракции, а 325 °С — верхнему температурному пределу фракции среднего масла. [c.185]
На диаграмме получают площади (их обычно заштриховывают), напоминающие по форме репу или луковицу и дающие наглядное представление о результатах разгонки. Дополнительно слева на симметричной диаграмме приводят ряд чисел, соответствующих количествам дистиллята (в %), которые были получены с момента начала разгонки до определенной температуры справа на диаграмме наносят числа, показывающие выход дистиллята (в %) для определенных температурных интервалов. На диаграмме разгонки (рис. 118, 6 значение 180 С соответствует верхнему температурному пределу бензиновой фракции, а 325 °С — верхнему температурному пределу фракции среднего масла. [c.185]
При газохроматографическом анализе смеси малеинимидов, полученных окислением деметаллированных порфиринов нефти и гилсонита, были количественно определены монометил-, моноэтил-метплзтил-, метилпропил-, этилпропилмалеинимиды. Ценность предложенной методики несколько снижают необходимость предварительного деметаллирования порфиринов (выход 60%) [832] и малая летучесть -малеинимидов, затрудняющая определение высших гомологов. Для определения структуры малеинимидов мы применяли исчерпывающее аналитическое гидрирование в токе водорода на платиновом катализаторе, помещенном в испаритель [c.150]
Для определения структуры малеинимидов мы применяли исчерпывающее аналитическое гидрирование в токе водорода на платиновом катализаторе, помещенном в испаритель [c.150]
Недавно предложен метод определения открытых положений в ископаемых порфиринах, основанный на реакции электрофиль-ного замещения пиррольных атомов водорода в молекулах порфиринов на атомы брома [833]. На синтетических ванадилпорфириновых комплексах показано, что реакция проходит исчерпывающе и достаточно селективно. По разработанной методике проведено бромирование порфириновых концентратов нефтей Западной Сибири и Южного Узбекистана. Состав продуктов [357] селективного бромирования установлен методом фракционной разгонки в масс-спектрометре по полному ионному току. При этом установлено, что молекулы ванадилпорфиринов нефтей содержат от одного да трех открытых положений на пиррольных кольцах, причем относительное содержание таких соединений достигает 70% общего количества нефтяных ванадилпорфиринов и меняется для ра лич-ных нефтей. Распределение порфиринов, содержащих одно и два незамещенных пиррольных положения в молекуле, для гомологов ряда М одной из нефтей Западной Сибири приведено на рис. 5.2. Несколько неожиданным оказалось, что пиррольные протоны характерны нё только для низкомолекулярных ванадилпорфиринов. [c.151]
Распределение порфиринов, содержащих одно и два незамещенных пиррольных положения в молекуле, для гомологов ряда М одной из нефтей Западной Сибири приведено на рис. 5.2. Несколько неожиданным оказалось, что пиррольные протоны характерны нё только для низкомолекулярных ванадилпорфиринов. [c.151]
Этот метод может быть распространен и на определенные виды аналогичных реакций гомологов, обладающих сходной структурной формулой (например, 2-метилалканов). Метод применим и к таким гомологам и к аналогичным реакциям как в форме сопоставления разностей, так и в форме сопоставления отнощений (см. 18, 23 и 24). Впрочем, и допущение о постоянстве разностей, и допущение о постоянстве отношений должны рассматриваться как упрощение метода, дающее возможность ограничиться для исследуемых реакций или соединений исходными данными для одной температуры, в частности для 298 К. Более общей формой является линейная зависимость между сопоставляемыми величинами, которая в частных случаях приводит к постоянству разностей или отношений. [c.279]
[c.279]
Для определения температурной зависимости параметров реакций образования гомологов простое сравнение в той или другой степени применимо только для расчета теплот образования и связанных с ней величин, но не для A5f, АС/ и Igi f. При расчете AHj можно пользоваться и допущением о постоянстве разностей, и допущением о постоянстве отношений. Оба они дают в этом случае довольно значительную погрешность, но первый путь обычно несколько точнее, и в дальнейшем здесь можно ограничиться в основном иллюстрацией метода разностей. В табл. VH, 13 сопоставлены теплоты образования AHf алканов. Изменение разностей и Яг при изменении температуры от 298 до 1500 К достигает, например, для -бутана — к-пентана 0,9 при изменении AH°f [c.287]
В связи с наличием в молекуле двухатомного фенола двух подвижных атомов водорода имеется возможность лучше подобрать величины гидрофобной и гидрофильной частей и соотношение между ними для получения наиболее эффективного деэмульгатора. После присоединения к двухатомному фенолу цепи окиси пропилена в полученной гидрофобной части имеется две концевые гидроксильные группы, к которым можно присоединить две гидрофильные цепи окиси этилена. Можно было ожидать, что на основе соединений подобного строения будут получены высокоэффективные деэмульгаторы. Были синтезированы блоксополимеры с цепями окисей пропилена и этилена различной длины на основе пирокатехина, резорцина и их гомологов. При этом к двухатомному фенолу присоединяли сначала цень окиси иронилена определенной длины, а затем цепи окиси этилена. [c.132]
После присоединения к двухатомному фенолу цепи окиси пропилена в полученной гидрофобной части имеется две концевые гидроксильные группы, к которым можно присоединить две гидрофильные цепи окиси этилена. Можно было ожидать, что на основе соединений подобного строения будут получены высокоэффективные деэмульгаторы. Были синтезированы блоксополимеры с цепями окисей пропилена и этилена различной длины на основе пирокатехина, резорцина и их гомологов. При этом к двухатомному фенолу присоединяли сначала цень окиси иронилена определенной длины, а затем цепи окиси этилена. [c.132]
Какие же структуры разветвленных алканов можно отнести к углеводородам изопреноидного типа строения Строго говоря, терминология здесь несколько произвольна, так как изопреноидные алканы нефтей не обязательно состоят из, отдельных изопреновых единиц. В этих углеводородах, как в типичных реликтах, проявляется их гомологичность и, конечно, неравновесность . Критерием для отнесения алканов к изопреноидным углеводородам служит правильное чередование метильных групп. Гомологичность является, как и всюду, следствием процессов деструкции более высокомолекулярных источников. Однако в отличие от реликтовых не-разветвленных алканов в изонреноидах всегда можно обнаружить провалы в концентрациях тех или иных гомологов. Эти провалы (отсутствие или малые относительные концентрации) некоторых гомологов являются следствием невозможности разрыва цепи (образования гомолога) в том месте, где находятся замещающие ме-тильные радикалы. Эта особенность чрезвычайно важна для определения источников образования тех или иных изопреноидных алканов. Именно отсутствие некоторых гомологов дает иногда наиболее ценную информацию. [c.60]
Гомологичность является, как и всюду, следствием процессов деструкции более высокомолекулярных источников. Однако в отличие от реликтовых не-разветвленных алканов в изонреноидах всегда можно обнаружить провалы в концентрациях тех или иных гомологов. Эти провалы (отсутствие или малые относительные концентрации) некоторых гомологов являются следствием невозможности разрыва цепи (образования гомолога) в том месте, где находятся замещающие ме-тильные радикалы. Эта особенность чрезвычайно важна для определения источников образования тех или иных изопреноидных алканов. Именно отсутствие некоторых гомологов дает иногда наиболее ценную информацию. [c.60]
Качественное и количественное определение в нефтях (фракция 200—250° С) адамантанов стало значительно более надежным после разработки специального метода — гидрокрекинга, в процессе которого углеводороды неадамантановой структуры претерпевают деструкцию, в то время как адамантан и его гомологи остаются неизмененными [18]. [c.102]
[c.102]
Впервые реакция метиленирования насыщенных углеводородов была описана Дёрингом и Баттери [118]. В результате реакции получается смесь специфических (т. е. некоторых, строго определенных) изомеров, являющихся очередными гомологами исходного углеводорода. [c.291]
Ожидаемый состав продуктов метиленирования удобно представлять в виде частей образующихся углеводородов. Количество этих частей соответствует количеству связей С—Н определенного типа, образующих при метиленировании тот или иной гомолог. Понятно, что общая сумма образовавшихся частей должна быть равна числу атомов водорода в исходном углеводороде. Например, при метиленировании этилциклопентапа (С7Н14) можно было ожидать образование одной части 1-метил-1-этилциклопен-тана, трех частей пропилциклопентана и двух частей для каждого из остальных пяти образующихся изомеров, т. е. всего 14 частей. Данные табл. 77, а также хроматограмма продуктов метилепиро- [c.293]
В технологических процессах, связанных с получением, переработкой и транспортированием горючих газов и паров, всегда имеется опасноспь существования взрывчатых паро-газовых систем. Так, взрывоопасные смеси могут образовываться при утечке горючих газов в атмосферу, при подсосе атмосферного воздуха в вакуумиро-ванные аппараты либо при неправильной работе технологических агрегатов, вследствие которой газовые потоки направляются в линии, для них не предназначенные. Многие технологические процессы связаны с проведением реакций между компонентами, смеси которых взрывчаты в определенном диапазоне составов. В ряде случаев регламент процесса предусматривает образование горючей смеси, например при окислительном пиролизе углеводородов. Наконец, ряд многотонпажных производств связан с синтезированием и переработкой продуктов, способных к взрывному распаду ацетилена и его гомологов, окиси этилена, закиси азота, озона, перекиси водорода и других. [c.60]
Так, взрывоопасные смеси могут образовываться при утечке горючих газов в атмосферу, при подсосе атмосферного воздуха в вакуумиро-ванные аппараты либо при неправильной работе технологических агрегатов, вследствие которой газовые потоки направляются в линии, для них не предназначенные. Многие технологические процессы связаны с проведением реакций между компонентами, смеси которых взрывчаты в определенном диапазоне составов. В ряде случаев регламент процесса предусматривает образование горючей смеси, например при окислительном пиролизе углеводородов. Наконец, ряд многотонпажных производств связан с синтезированием и переработкой продуктов, способных к взрывному распаду ацетилена и его гомологов, окиси этилена, закиси азота, озона, перекиси водорода и других. [c.60]
Определение производится следующим образом. В мерный цилиндр с притертой пробкой емкостью 100 мл наливают 25 мл раствора изатина в серной кислоте (раствор изатина готовят растворением 0,5 г изатина в 1000 г концентрированной серной кислоты), затем добавляют 2 мл исследуемого продукта и 25 мл концентрированной сер1[ой кислоты, к которой заранее добавлена 1 капля концентрированной азотной кислоты. При наличии тиофена или его гомологов появляется соответствующее окрашивание. [c.391]
При наличии тиофена или его гомологов появляется соответствующее окрашивание. [c.391]
| 1. | Пара гомологов | 1 вид — рецептивный | лёгкое | 1 Б. | Знать определение и примеры гомологов метана. |
| 2. | Характерные признаки алканов | 1 вид — рецептивный | лёгкое |
1 Б.
|
Знать общие признаки гомологического ряда метана. |
| 3. | Формулы и названия алканов | 1 вид — рецептивный | лёгкое | 1 Б. | Знать формулы и названия алканов. |
| 4. | Отличия гомологов | 1 вид — рецептивный | лёгкое |
2 Б.
|
Знать примеры и отличительные особенности гомологов. |
| 5. | Состав и строение | 2 вид — интерпретация | среднее | 2 Б. | Узнавать признаки алканов, изображённых на рисунке. |
| 6. | Заполни пропуски | 2 вид — интерпретация | среднее |
2 Б.
|
Знать гомологический ряд предельных углеводородов алканов. Уметь сравнивать и обобщать. |
| 7. | Состав гомологов метана | 2 вид — интерпретация | среднее | 2 Б. | Знать состав молекул гомологов метана, общую формулу гомологического ряда алканов. |
8.
|
Выбери верный ряд | 3 вид — анализ | сложное | 3 Б. | Знать гомологический ряд алканов, общую формулу алканов, гомологическую разность, уметь анализировать информацию. |
| 9. | Молярная масса гомологов метана | 3 вид — анализ | сложное |
3 Б.
|
Знать состав молекул гомологов метана, общую формулу гомологического ряда алканов, уметь рассчитывать молярную массу, применять знание закономерностей изменения молярной массы веществ в гомологическом ряду. |
| 10. | Подбери значение | 3 вид — анализ | сложное | 3 Б. |
Знать формулы алканов, общую формулу гомологического ряда алканов, уметь применять знание закономерностей изменения молярной массы веществ в гомологическом ряду и понятие гомологической разности. Уметь проводить расчёты, сравнивать и обобщать. Уметь проводить расчёты, сравнивать и обобщать.
|
Гомологи
1
H
1,008
1s1
2,2
Бесцветный газ
t°пл=-259°C
t°кип=-253°C
2
He
4,0026
1s2
Бесцветный газ
t°кип=-269°C
3
Li
6,941
2s1
0,99
Мягкий серебристо-белый металл
t°пл=180°C
t°кип=1317°C
4
Be
9,0122
2s2
1,57
Светло-серый металл
t°пл=1278°C
t°кип=2970°C
5
B
10,811
2s2 2p1
2,04
Темно-коричневое аморфное вещество
t°пл=2300°C
t°кип=2550°C
6
C
12,011
2s2 2p2
2,55
Прозрачный (алмаз) / черный (графит) минерал
t°пл=3550°C
t°кип=4830°C
7
N
14,007
2s2 2p3
3,04
Бесцветный газ
t°пл=-210°C
t°кип=-196°C
8
O
15,999
2s2 2p4
3,44
Бесцветный газ
t°пл=-218°C
t°кип=-183°C
9
F
18,998
2s2 2p5
4,0
Бледно-желтый газ
t°пл=-220°C
t°кип=-188°C
10
Ne
20,180
2s2 2p6
Бесцветный газ
t°пл=-249°C
t°кип=-246°C
11
Na
22,990
3s1
0,93
Мягкий серебристо-белый металл
t°пл=98°C
t°кип=892°C
12
Mg
24,305
3s2
1,31
Серебристо-белый металл
t°пл=649°C
t°кип=1107°C
13
Al
26,982
3s2 3p1
1,61
Серебристо-белый металл
t°пл=660°C
t°кип=2467°C
14
Si
28,086
3s2 3p2
1,9
Коричневый порошок / минерал
t°пл=1410°C
t°кип=2355°C
15
P
30,974
3s2 3p3
2,2
Белый минерал / красный порошок
t°пл=44°C
t°кип=280°C
16
S
32,065
3s2 3p4
2,58
Светло-желтый порошок
t°пл=113°C
t°кип=445°C
17
Cl
35,453
3s2 3p5
3,16
Желтовато-зеленый газ
t°пл=-101°C
t°кип=-35°C
18
Ar
39,948
3s2 3p6
Бесцветный газ
t°пл=-189°C
t°кип=-186°C
19
K
39,098
4s1
0,82
Мягкий серебристо-белый металл
t°пл=64°C
t°кип=774°C
20
Ca
40,078
4s2
1,0
Серебристо-белый металл
t°пл=839°C
t°кип=1487°C
21
Sc
44,956
3d1 4s2
1,36
Серебристый металл с желтым отливом
t°пл=1539°C
t°кип=2832°C
22
Ti
47,867
3d2 4s2
1,54
Серебристо-белый металл
t°пл=1660°C
t°кип=3260°C
23
V
50,942
3d3 4s2
1,63
Серебристо-белый металл
t°пл=1890°C
t°кип=3380°C
24
Cr
51,996
3d5 4s1
1,66
Голубовато-белый металл
t°пл=1857°C
t°кип=2482°C
25
Mn
54,938
3d5 4s2
1,55
Хрупкий серебристо-белый металл
t°пл=1244°C
t°кип=2097°C
26
Fe
55,845
3d6 4s2
1,83
Серебристо-белый металл
t°пл=1535°C
t°кип=2750°C
27
Co
58,933
3d7 4s2
1,88
Серебристо-белый металл
t°пл=1495°C
t°кип=2870°C
28
Ni
58,693
3d8 4s2
1,91
Серебристо-белый металл
t°пл=1453°C
t°кип=2732°C
29
Cu
63,546
3d10 4s1
1,9
Золотисто-розовый металл
t°пл=1084°C
t°кип=2595°C
30
Zn
65,409
3d10 4s2
1,65
Голубовато-белый металл
t°пл=420°C
t°кип=907°C
31
Ga
69,723
4s2 4p1
1,81
Белый металл с голубоватым оттенком
t°пл=30°C
t°кип=2403°C
32
Ge
72,64
4s2 4p2
2,0
Светло-серый полуметалл
t°пл=937°C
t°кип=2830°C
33
As
74,922
4s2 4p3
2,18
Зеленоватый полуметалл
t°субл=613°C
(сублимация)
34
Se
78,96
4s2 4p4
2,55
Хрупкий черный минерал
t°пл=217°C
t°кип=685°C
35
Br
79,904
4s2 4p5
2,96
Красно-бурая едкая жидкость
t°пл=-7°C
t°кип=59°C
36
Kr
83,798
4s2 4p6
3,0
Бесцветный газ
t°пл=-157°C
t°кип=-152°C
37
Rb
85,468
5s1
0,82
Серебристо-белый металл
t°пл=39°C
t°кип=688°C
38
Sr
87,62
5s2
0,95
Серебристо-белый металл
t°пл=769°C
t°кип=1384°C
39
Y
88,906
4d1 5s2
1,22
Серебристо-белый металл
t°пл=1523°C
t°кип=3337°C
40
Zr
91,224
4d2 5s2
1,33
Серебристо-белый металл
t°пл=1852°C
t°кип=4377°C
41
Nb
92,906
4d4 5s1
1,6
Блестящий серебристый металл
t°пл=2468°C
t°кип=4927°C
42
Mo
95,94
4d5 5s1
2,16
Блестящий серебристый металл
t°пл=2617°C
t°кип=5560°C
43
Tc
98,906
4d6 5s1
1,9
Синтетический радиоактивный металл
t°пл=2172°C
t°кип=5030°C
44
Ru
101,07
4d7 5s1
2,2
Серебристо-белый металл
t°пл=2310°C
t°кип=3900°C
45
Rh
102,91
4d8 5s1
2,28
Серебристо-белый металл
t°пл=1966°C
t°кип=3727°C
46
Pd
106,42
4d10
2,2
Мягкий серебристо-белый металл
t°пл=1552°C
t°кип=3140°C
47
Ag
107,87
4d10 5s1
1,93
Серебристо-белый металл
t°пл=962°C
t°кип=2212°C
48
Cd
112,41
4d10 5s2
1,69
Серебристо-серый металл
t°пл=321°C
t°кип=765°C
49
In
114,82
5s2 5p1
1,78
Мягкий серебристо-белый металл
t°пл=156°C
t°кип=2080°C
50
Sn
118,71
5s2 5p2
1,96
Мягкий серебристо-белый металл
t°пл=232°C
t°кип=2270°C
51
Sb
121,76
5s2 5p3
2,05
Серебристо-белый полуметалл
t°пл=631°C
t°кип=1750°C
52
Te
127,60
5s2 5p4
2,1
Серебристый блестящий полуметалл
t°пл=450°C
t°кип=990°C
53
I
126,90
5s2 5p5
2,66
Черно-серые кристаллы
t°пл=114°C
t°кип=184°C
54
Xe
131,29
5s2 5p6
2,6
Бесцветный газ
t°пл=-112°C
t°кип=-107°C
55
Cs
132,91
6s1
0,79
Мягкий серебристо-желтый металл
t°пл=28°C
t°кип=690°C
56
Ba
137,33
6s2
0,89
Серебристо-белый металл
t°пл=725°C
t°кип=1640°C
57
La
138,91
5d1 6s2
1,1
Серебристый металл
t°пл=920°C
t°кип=3454°C
58
Ce
140,12
f-элемент
Серебристый металл
t°пл=798°C
t°кип=3257°C
59
Pr
140,91
f-элемент
Серебристый металл
t°пл=931°C
t°кип=3212°C
60
Nd
144,24
f-элемент
Серебристый металл
t°пл=1010°C
t°кип=3127°C
61
Pm
146,92
f-элемент
Светло-серый радиоактивный металл
t°пл=1080°C
t°кип=2730°C
62
Sm
150,36
f-элемент
Серебристый металл
t°пл=1072°C
t°кип=1778°C
63
Eu
151,96
f-элемент
Серебристый металл
t°пл=822°C
t°кип=1597°C
64
Gd
157,25
f-элемент
Серебристый металл
t°пл=1311°C
t°кип=3233°C
65
Tb
158,93
f-элемент
Серебристый металл
t°пл=1360°C
t°кип=3041°C
66
Dy
162,50
f-элемент
Серебристый металл
t°пл=1409°C
t°кип=2335°C
67
Ho
164,93
f-элемент
Серебристый металл
t°пл=1470°C
t°кип=2720°C
68
Er
167,26
f-элемент
Серебристый металл
t°пл=1522°C
t°кип=2510°C
69
Tm
168,93
f-элемент
Серебристый металл
t°пл=1545°C
t°кип=1727°C
70
Yb
173,04
f-элемент
Серебристый металл
t°пл=824°C
t°кип=1193°C
71
Lu
174,96
f-элемент
Серебристый металл
t°пл=1656°C
t°кип=3315°C
72
Hf
178,49
5d2 6s2
Серебристый металл
t°пл=2150°C
t°кип=5400°C
73
Ta
180,95
5d3 6s2
Серый металл
t°пл=2996°C
t°кип=5425°C
74
W
183,84
5d4 6s2
2,36
Серый металл
t°пл=3407°C
t°кип=5927°C
75
Re
186,21
5d5 6s2
Серебристо-белый металл
t°пл=3180°C
t°кип=5873°C
76
Os
190,23
5d6 6s2
Серебристый металл с голубоватым оттенком
t°пл=3045°C
t°кип=5027°C
77
Ir
192,22
5d7 6s2
Серебристый металл
t°пл=2410°C
t°кип=4130°C
78
Pt
195,08
5d9 6s1
2,28
Мягкий серебристо-белый металл
t°пл=1772°C
t°кип=3827°C
79
Au
196,97
5d10 6s1
2,54
Мягкий блестящий желтый металл
t°пл=1064°C
t°кип=2940°C
80
Hg
200,59
5d10 6s2
2,0
Жидкий серебристо-белый металл
t°пл=-39°C
t°кип=357°C
81
Tl
204,38
6s2 6p1
Серебристый металл
t°пл=304°C
t°кип=1457°C
82
Pb
207,2
6s2 6p2
2,33
Серый металл с синеватым оттенком
t°пл=328°C
t°кип=1740°C
83
Bi
208,98
6s2 6p3
Блестящий серебристый металл
t°пл=271°C
t°кип=1560°C
84
Po
208,98
6s2 6p4
Мягкий серебристо-белый металл
t°пл=254°C
t°кип=962°C
85
At
209,98
6s2 6p5
2,2
Нестабильный элемент, отсутствует в природе
t°пл=302°C
t°кип=337°C
86
Rn
222,02
6s2 6p6
2,2
Радиоактивный газ
t°пл=-71°C
t°кип=-62°C
87
Fr
223,02
7s1
0,7
Нестабильный элемент, отсутствует в природе
t°пл=27°C
t°кип=677°C
88
Ra
226,03
7s2
0,9
Серебристо-белый радиоактивный металл
t°пл=700°C
t°кип=1140°C
89
Ac
227,03
6d1 7s2
1,1
Серебристо-белый радиоактивный металл
t°пл=1047°C
t°кип=3197°C
90
Th
232,04
f-элемент
Серый мягкий металл
91
Pa
231,04
f-элемент
Серебристо-белый радиоактивный металл
92
U
238,03
f-элемент
1,38
Серебристо-белый металл
t°пл=1132°C
t°кип=3818°C
93
Np
237,05
f-элемент
Серебристо-белый радиоактивный металл
94
Pu
244,06
f-элемент
Серебристо-белый радиоактивный металл
95
Am
243,06
f-элемент
Серебристо-белый радиоактивный металл
96
Cm
247,07
f-элемент
Серебристо-белый радиоактивный металл
97
Bk
247,07
f-элемент
Серебристо-белый радиоактивный металл
98
Cf
251,08
f-элемент
Нестабильный элемент, отсутствует в природе
99
Es
252,08
f-элемент
Нестабильный элемент, отсутствует в природе
100
Fm
257,10
f-элемент
Нестабильный элемент, отсутствует в природе
101
Md
258,10
f-элемент
Нестабильный элемент, отсутствует в природе
102
No
259,10
f-элемент
Нестабильный элемент, отсутствует в природе
103
Lr
266
f-элемент
Нестабильный элемент, отсутствует в природе
104
Rf
267
6d2 7s2
Нестабильный элемент, отсутствует в природе
105
Db
268
6d3 7s2
Нестабильный элемент, отсутствует в природе
106
Sg
269
6d4 7s2
Нестабильный элемент, отсутствует в природе
107
Bh
270
6d5 7s2
Нестабильный элемент, отсутствует в природе
108
Hs
277
6d6 7s2
Нестабильный элемент, отсутствует в природе
109
Mt
278
6d7 7s2
Нестабильный элемент, отсутствует в природе
110
Ds
281
6d9 7s1
Нестабильный элемент, отсутствует в природе
Металлы
Неметаллы
Щелочные
Щелоч-зем
Благородные
Галогены
Халькогены
Полуметаллы
s-элементы
p-элементы
d-элементы
f-элементы
Наведите курсор на ячейку элемента, чтобы получить его краткое описание.
Чтобы получить подробное описание элемента, кликните по его названию.
Гомологи метана — насыщенные углеводороды
Химия. 9 класс. Григорович
Вспомните:
- молекулярные формулы отражают только качественный и количественный состав веществ, а структурные — еще и порядок соединения атомов;
- при составлении структурных формул каждую химическую связь между атомами обозначают черточкой.
Гомологи и гомологический ряд метана
В предыдущем параграфе вы уже ознакомились с простейшим углеводородом — метаном, являющимся также простейшим представителем класса органических соединений алканов. Алканы — достаточно большой класс соединений, но у всех представителей алканов есть общий признак: их молекулярные формулы соответствуют общей формуле CnH2n+2, где n — число атомов Карбона в молекуле. Если вы вместо n подставите любое целое число, то получите молекулярную формулу алкана. Например, если подставить n = 1, то получим формулу метана CH4. Формулы и названия первых десяти представителей алканов представлены в таблице 10.
Например, если подставить n = 1, то получим формулу метана CH4. Формулы и названия первых десяти представителей алканов представлены в таблице 10.
Таблица 10. Первые десять представителей алканов
Число атомов Карбона n | Формула | Название |
1 | CH4 | Метан |
2 | C2H6 | Этан |
3 | C3H8 | Пропан |
4 | C4H10 | Бутан |
5 | C5H12 | Пентан |
Число атомов Карбона n | Формула | Название |
6 | C6H14 | Гексан |
7 | C7H16 | Гептан |
8 | C8H18 | Октан |
9 | C9H20 | Нонан |
10 | C10H22 | Декан |
Обратите внимание, что первые четыре представителя алканов — метан, этан, пропан и бутан — имеют исторически сложившиеся названия. Названия следующих алканов образуются от греческих числительных добавлением суффикса -ан-: пентан, гексан и т. д.
Названия следующих алканов образуются от греческих числительных добавлением суффикса -ан-: пентан, гексан и т. д.
Проанализируйте молекулярные формулы веществ в таблице. Как отличается состав молекул метана и этана? Формула метана — CH4, а этана — C2H6. Их формулы отличаются на один атом C и два атома H, т. е. на группу атомов CH2. Как отличается состав молекул этана и бутана? Они отличаются на два атома C и четыре атома H, т. е. на две группы атомов CH2.
• Соединения одного класса, имеющие сходное строение, но отличающиеся по составу на одну или несколько групп CH2, называют гомологами.
Группу CH2 называют гомологической разницей. Совокупность всех гомологов образует гомологический ряд.
Метан и его гомологи образуют гомологический ряд класса алканов. Первые представители этого ряда с небольшим числом атомов Карбона в молекуле называют низшими алканами, а с большим числом — высшими. Впрочем, четкой границы между ними нет.
Впрочем, четкой границы между ними нет.
Структурные формулы и строение молекул гомологов метана
Еще в начале развития органической химии ученые выяснили, что подавляющее большинство органических соединений являются веществами молекулярного строения. Но при определении формул органических веществ возникла проблема, связанная с валентностью элементов в их составе. К примеру, если определять валентность атомов Карбона в этане C2H6, то по правилам, которые вы изучали в 7 классе, у Карбона должна быть валентность III, что противоречит действительности. Исследования Ф. Кекуле и А. Бутлерова доказали, что формально вопрос валентности в органической химии рассматривать нельзя. Валентность Карбона в органических соединениях всегда IV, это становится понятным, если вместо молекулярных формул использовать структурные. Именно поэтому в органической химии используют структурные формулы, а молекулярные формулы — для решения расчетных задач, поскольку по ним легче рассчитывать молярную массу.
При составлении структурных формул следует помнить, что атомы Карбона в молекулах органических веществ соединены друг с другом и образуют карбоновую цепь (карбоновый скелет).
Рассмотрим составление структурных формул на примере этана.
1. Изображаем карбоновую цепь — в этане она состоит из двух атомов Карбона, соединенных одинарной связью:
C-C
2. Карбон четырехвалентен, поэтому от каждого атома Карбона рисуем черточки так, чтобы у каждого атома их было по четыре:
3. Дописываем символы атомов Гидрогена и получаем структурную формулу этана:
Часто структурные формулы записывают в сокращенном виде, не изображая связи C-H. Сокращенные структурные формулы намного компактнее, чем развернутые:
CH3-CH3
Структурные формулы показывают последовательность соединения атомов, но не отражают пространственного строения молекул. Атомы Карбона в молекулах гомологов метана находятся в возбужденном состоянии, как и в молекулах метана, о чем вы узнали в предыдущем параграфе. Значит, все химические связи от каждого атома Карбона направлены к вершинам тетраэдра, и молекула этана имеет такой пространственный вид:
Значит, все химические связи от каждого атома Карбона направлены к вершинам тетраэдра, и молекула этана имеет такой пространственный вид:
По приведенному алгоритму можно составить структурные формулы других углеводородов:
При составлении структурных формул соединений других классов также следует соблюдать эти принципы и, главное, помнить и правильно учитывать валентность элементов.
Проанализируйте все структурные формулы, приведенные в этом разделе. Видно, что атомы Карбона четырехвалентны, к тому же, каждый атом Карбона соединяется с четырьмя другими атомами (Карбона или Гидрогена). Значит, все валентные возможности атомов Карбона «насыщены» другими атомами, и они уже не способны присоединить ни одного атома. Такие углеводороды называют насыщенными. Главным признаком насыщенных соединений является то, что в их молекулах все связи между атомами Карбона одинарные.
• Насыщенные соединения — это органические соединения, в молекулах которых атомы Карбона соединены друг с другом только одинарными ковалентными связями.
Метан и его гомологи (углеводороды класса алканов) — это насыщенные углеводороды.
• Название первого члена гомологического ряда алканов «метан» произошло от названия соответствующего ему спирта — метилового. Этот спирт издавна называли древесным, поскольку добывали его из древесины. Слово «метил» происходит от греческих слов methy — вино и hile — лес (буквально — «вино из леса»).
• Алкан с самой длинной карбоновой цепью — нонаконтатриктан С390Н782— синтезировали в 1985 году английские химики И. Бидд и М. Уайтинг.
Ключевая идея
Для каждого класса органических соединений характерен свой гомологический ряд. Состав соединений одного ряда отражает общая формула.
Контрольные вопросы
- 270. Какова валентность атомов Карбона в органических соединениях?
- 271. Какие углеводороды называют алканами? Приведите их общую формулу.
- 272. Почему метан и его гомологи называют насыщенными углеводородами?
- 273.
 Дайте определение понятиям «гомолог», «гомологический ряд», «гомологическая разница».
Дайте определение понятиям «гомолог», «гомологический ряд», «гомологическая разница». - 274. Что общего и различного в строении двух гомологов?
 Дайте определение понятиям «гомолог», «гомологический ряд», «гомологическая разница».
Дайте определение понятиям «гомолог», «гомологический ряд», «гомологическая разница».Задания для усвоения материала
275. Составьте полную и сокращенную структурные формулы гомологов метана с числом атомов Карбона 6 и 8.
276. Докажите, что пропан и октан являются гомологами.
277. Сколько химических связей в молекуле пропана? Сколько в них связей C-C и сколько C-H?
278. Из приведенного перечня формул углеводородов выпишите формулы гомологов метана: C3H6, C4H10, C6H6, C14H30, C8H8.
279. Определите массовые доли Карбона в метане и пропане. В каком веществе массовая доля Карбона больше? Можно ли из этого сделать вывод о том, как меняется массовая доля Карбона в алканах с увеличением числа атомов Карбона в молекуле? Можно ли сделать такой же вывод без расчета массовых долей?
280. Какой гомолог метана обладает плотностью, почти равной плотности воздуха?
Какой гомолог метана обладает плотностью, почти равной плотности воздуха?
Гомология
Гомологи метана. Скорость взаимодействия с гидроксильным радикалом сильно зависит от строения молекулы алкана. Вторичные и третичные атомы водорода метиленовых (—СН2—) и метино-вых ( —СН ) групп отщепляются легче, чем первичные.[ …]
Гомологи фенола, замещенные в пара-положении, с п-нитроани-лином не определяются.[ …]
Типовая тетаграмма (кривая распределения эквивалентно-потенциальной температуры с высотой), характерная для определенного географического типа воздушных масс в данном месте.[ …]
Содержание в стоках гомологов бензола С7—С10 с одним, двумя, тремя и четырьмя заместителями в ядре увеличивается при переработке смешанных и ароматических нефтей. Заместителями в ядре чаще всего являются метильные радикалы, т. е. основная масса моноциклических ароматических углеводородов в стоках представлена полиметилбензолами.[ …]
Вещество, соответствующее пику (б), и его гомологи были идентифицированы с помощью сочетания ГХ/МС, как серия этоксикарбинолов (рис.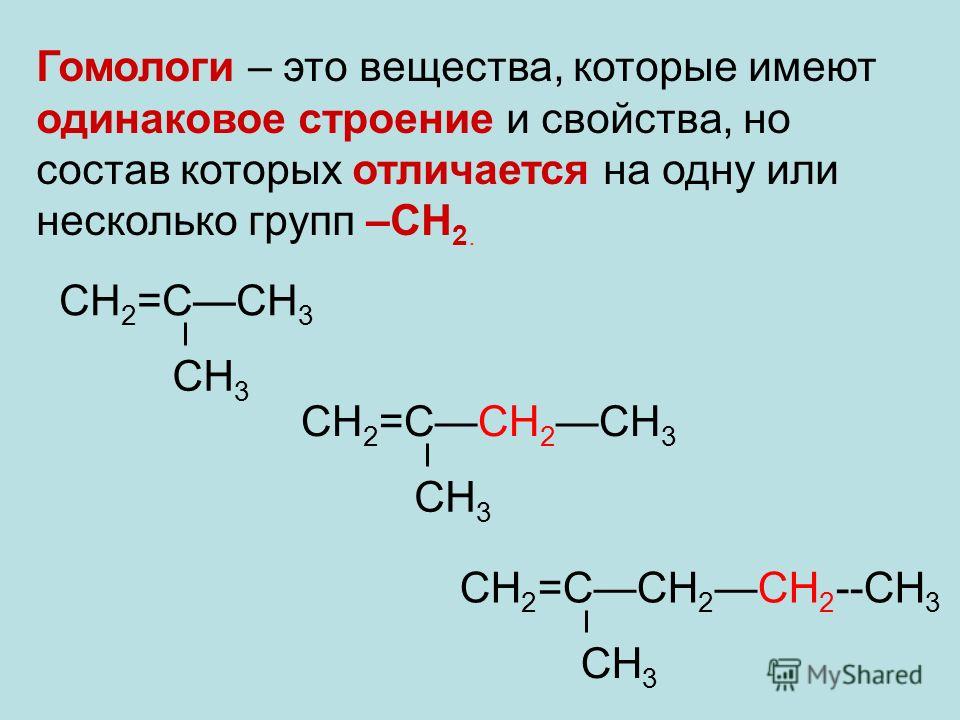 Элементный состав и наличие атомов кислорода были ясно продемонстрированы результатами АЭД-анализа (рис. Однако структура, предложенная в результате библиотечного поиска, масс-спектров и ИК-спектров, оказалась неверной. Для окончательного выяснения структуры было бы необходимо провести хромато-масс-спектрометриче-ский анализ в режиме химической ионизации для регистрации молекулярного иона (см. раздел 1.2), но при решении поставленной задачи достаточно было показать, что данное соединение является неионогенным поверхностно-активным веществом (ПАВ).[ …]
Элементный состав и наличие атомов кислорода были ясно продемонстрированы результатами АЭД-анализа (рис. Однако структура, предложенная в результате библиотечного поиска, масс-спектров и ИК-спектров, оказалась неверной. Для окончательного выяснения структуры было бы необходимо провести хромато-масс-спектрометриче-ский анализ в режиме химической ионизации для регистрации молекулярного иона (см. раздел 1.2), но при решении поставленной задачи достаточно было показать, что данное соединение является неионогенным поверхностно-активным веществом (ПАВ).[ …]
Вместе с тем продолжается дискуссия о физиологической гомологии добавочных желез сомовидных рыб с семенными пузырьками млекопитающих, у которых последние, как известно, являются местом хранения спермы и вырабатывают секрет, продлевающий срок жизни половых клеток.[ …]
Кубовый остаток представляет собой высококипящие эфиры высших гомологов уксусной кислоты (пропионовой, масляной к других кислот). Этот остаток спускают из куба в монжус лишь после трех оборотов установки. Из монжуса высококипящие эфиры перепускают в перегонный куб для отгонки из них смеси сложных эфиров: бутилацетата, бутилпропионата, бутилбутирата и др. Остаток после отгонки спускают в канализацию.[ …]
Из монжуса высококипящие эфиры перепускают в перегонный куб для отгонки из них смеси сложных эфиров: бутилацетата, бутилпропионата, бутилбутирата и др. Остаток после отгонки спускают в канализацию.[ …]
По свойствам синтетическое вещество было идентично природному. Гомолог этого вещества—транс-октен-2-ол-1-ацетат-1, выделенный у клопа—бронзовки (Rhaecocoris subciventris Stal.), по-видимому, является половым атрактантом этих насекомых1.[ …]
Возможно этот фактор оказался решающим в реализации опережающей экспрессии модифицирующего компонента при установлении клонированных генов RMBsuR I в E. coli.[ …]
Другим, может быть и менее неожиданным, результатом сравнения является отсутствие заметной гомологии среди рестриктаз [109,199,382], даже между ферментами с близкими характеристиками субстратной специфичности как, например, НРаеИ7 I (5’С ТСОАО) и I (5’ ТСОА) [350]. В то же время иммунологическими методами обнаружено сходство изошизомеров Ивг I и ЕсоИ I на уровне третичной структуры [147]. [ …]
[ …]
В залежи содержится в свободном газообразном состоянии преимущественно метан; более тяжелые гомологи метана поглощаются нефтью. Поэтому среди углеводородных газов предельные преобладают над непредельными. Из прочих газов присутствует углекислый газ, растворимость которого в нефти больше, чем в воде, поэтому, помимо содержания СОг в газообразном состоянии, фиксируется его растворенная форма в нефти. Азот же в нефти растворяется плохо, поэтому он находится преимущественно в свободной газообразной фазе.[ …]
В выхлопных газах автомобильных двигателей присутствуют до 150 ПАУ, их замещенных производных и гомологов [50,51].[ …]
Ни для одной вирусной РНК пока не удалось определить полную нуклеотидную последовательность. Однако степень гомологии между вирусами можно было бы определять при помощи гибридизации с использованием двухцепочечпых форм РНК вирусов растений; этот метод вполне может оказаться пригодным как для распределения родственных вирусов по группам, так и для оценки степени родства в каждой группе. Другой способ установления степени родства состоит в определении частоты 16 возможных динуклеотидов в различных нуклеиновых кислотах. Этот прием до сих пор использовался только в работе с ДНК-содержащими вирусами [1694].[ …]
Другой способ установления степени родства состоит в определении частоты 16 возможных динуклеотидов в различных нуклеиновых кислотах. Этот прием до сих пор использовался только в работе с ДНК-содержащими вирусами [1694].[ …]
Органические кислоты на 2 /а состоят из кислот, содержащих 1—2 углеродных атома, однако среди них имеются также гомологи с 3—6 углеродными атомами [12].[ …]
Степень извлечения алкилсульфатов натрия из водных растворов увеличивается гари переходе от низшего к более высшему гомологу данного ряда.[ …]
Простейший циклоалкан — циклопропан — в организме не разрушается и выделяется в неизмененном виде с выдыхаемым воздухом и через кожу. Другие гомологи депонируются в тканях и подвергаются биотрансформации, причем по мере увеличения размеров молекул снижается количество метаболитов.[ …]
В настоящее время за рубежом и в нашей стране применяется новый метод определения фенолов с 4-аминоантипирином. Он основан на том, что фенол и его гомологи в щелочной среде в присутствии окислителя образуют окрашенное соединение с 4-аминоантипирином, так называемый красный антипирин. [ …]
[ …]
В верху абсорбционной колонны поддерживается температура в 106—107°. Из нижней части ее в обезвоживающую колонну 26, вместе с маслами уходит уксусная кислота, ее гомологи и немного воды. Вода и часть уксусной кислоты в виде паров уходят из обезвоживающей колонны 26 обратно в абсорбционную колонну 18. Из нижней части обезвоживающей колонны уксусная кислота и ее гомологи, смоляные масла и небольшое количество воды переходят в вакуум-колонну 27, где кислоту и воду отгоняют, а масла из нижней части этой колонны уходят в холодильник 28, из которого их насосом 29 подают в бак 30 для смоляных масел, из него в бак 30а и далее насосом 31 или 32 — в напорный бак 19 при абсорбционной колонне. Таким образом, смоляные масла являются оборотным материалом.[ …]
Циклоалканы по физико-химическим свойствам очень сходны с соответствующими алканами. Агрегатное состояние — бесцветные газы (циклопропан) или жидкости, высшие гомологи -твердые вещества. Трех и четырехчленные кольца менее устойчивы: циклопропан легко гидролизуется при 120 °С, циклобутан — при 180 °С. Циклопентановые производные гидрируются в более жестких условиях — при 300°С в присутствии катализаторов. Циклогексан вообще не подвергается гидрированию.[ …]
Циклопентановые производные гидрируются в более жестких условиях — при 300°С в присутствии катализаторов. Циклогексан вообще не подвергается гидрированию.[ …]
Арены содержатся в нефтях от 10 до 20 %, редко достигая 35 % и более. Наиболее богаты аренами молодые кайнозойские нефти. Этот класс углеводородов представлен в нефтях гомологами бензола, производными би- и полициклических соединений. В нефтях идентифицированы гомологи нафталина, дифенила.[ …]
Осмотры 1 раз в 12 месяцев при анилиновом крашении на текстильных фабриках и крашении мехов урсоло-выми красителями. В случае возможности контакта с канцерогенными аминами— цистоскопия при поступлении на работу [125]. Отстранение от работы лиц с заболеваниями мочевых путей. Медицинское наблюдение за лицами, переведенными на работу вне контакта с ароматическими аминами или оставившими работу на данном предприятии. Темкин [125] считает целесообразным в анилинокрасочной промышленности принимать на работу лиц старшего возраста (меньше шансов на развитие рака, чем у молодых людей) и не рекомендует ограничивать срок работы в ней (см. также работу Карапетяна и Темкина).[ …]
также работу Карапетяна и Темкина).[ …]
Спирты получаются брожением крахмалистых веществ (пшеницы, картофеля). При разгонке перебродившей массы отгоняется главным образом этиловый спирт, а затем смесь высших гомологов — сивушные масла.[ …]
О фитогенной природе этих соединений говорит особенность их распределения внутри гомологических рядов. Так, в случае н-алканов летнего аэрозоля преобладают нечетночисленные гомологи (С2, С23, С25 и т. д.), тогда как в случае высших спиртов и карбоновых кислот — четночисленные (рис. 4.6).[ …]
Растворимость нефти в различных газах существенно различается. Растворимость нефти в углекислом газе значительно выше, чем в метане. По данным Т. П. Жузе (1976 г.), добавка к метану его гомологов резко увеличивает растворяющую способность газовой смеси. Как и для водных растворов, фактор давления играет важную роль для выноса жидких УВ в виде газовых растворов. При температуре 200° С и давлении 88,2 МПа в 1 м3 сжатого газа может раствориться 169—212 кг нефти. При высоких температуре и давлении в сжатых газах могут практически раствориться все компоненты нефти (В. А. Соколов, 1965 г.). Указанное давление в поровом пространстве нефтематеринских пород формируется в интервале глубин 1,5—4 км, т. е. охватывают всю зону интенсивной генерации нефти.[ …]
При высоких температуре и давлении в сжатых газах могут практически раствориться все компоненты нефти (В. А. Соколов, 1965 г.). Указанное давление в поровом пространстве нефтематеринских пород формируется в интервале глубин 1,5—4 км, т. е. охватывают всю зону интенсивной генерации нефти.[ …]
Озон реагирует с бензолом. Пои действии озона на пиали.ц получается янтарный альдегид. Озон легко окисляет оле-фины. Образующиеся при этШг оответствующие озониды также разлагаются в воде и, как правило, при этом возникают альде-гиды и кетоны. Можно привести еще целый ряд примеров, когда при наличии кратных связей в результате присоединения озона к органическим веществам образуются озониды, однако в этом нет необходимости.[ …]
Исследования, посвященные определению первичной структуры генов рестрикции-модификации с самого начала не являлись самоцелью. Полученные данные послужили предпосылкой для поиска возможных гомологий структуры генов — наличия или отсутствия консервативных нуклеотидных (или выводимых из них аминоскислотных) последовательностей. Эти сведения являются необходимыми для решения таких вопросов как эволюция генов гш, определение участков белковых молекул ответственных за специфическое взаимодействие с субстратом и участвующих в формировании каталитического центра.[ …]
Эти сведения являются необходимыми для решения таких вопросов как эволюция генов гш, определение участков белковых молекул ответственных за специфическое взаимодействие с субстратом и участвующих в формировании каталитического центра.[ …]
Триниклические ароматические углеводороды — антрацен и фенантрен превращаются микроорганизмами способом, аналогичным микробиологической деградации нафталина.[ …]
В качестве модели целлюлозы (3-целлобиоза имеет недостаток, связанный со значительным влиянием «краевых эффектов», но полный рентгеноструктурный анализ более высоких членов гомологического ряда еще не выполнен. Рентгенограммы порошков [43, 80] гомологов выше целлотетраозы имеют сильное сходство с рентгенограммой целлюлозы II. ИК-спектры целлотетраозы и особенно целлопентаозы [81] подобны ИК-спектрам целлюлозы II (после дейтерирования менее упорядоченных областей) в области валентных колебаний О—Н-групп, что указывает на участие всех этих групп в водородных связях. Следует ожидать, что будут наблюдаться небольшие различия между четными и нечетными членами гомологического ряда в зависимости от того, являются соседними одни и те же или различные концевые звенья. Для целлотриозы были выращены очень тонкие монокристаллы, однако провести анализ дальше определения вероятной пространственной группы не удалось [70]. Данные по инфракрасным спектрам и рентгенограмма порошка указывают, что упаковка цепей в целлодекстринах, начиная с целлотетраозы, очень похожа на упаковку цепей в целлюлозе II, хотя период идентичности вдоль оси цепи зависит для низших гомологов от длины молекулы [71]. Анализ монокристаллов целлотетраозы и целлопентаозы может дать очень интересные данные, однако он очень утомителен. Согласно сообщению [41] об изучении двумерной проекции (х, г; размеры осей такие же, как, указаны в табл. 1.13), кристаллы целлотетраозы имеют триклинную элементарную ячейку, длина которой в два раза больше длины ангидроцеллобиозного звена макромолекулы. Эти предварительные данные для монокристалла могут служить подтверждением того, что элементарная ячейка целлюлозы II является триклинной и содержит параллельно ориентированные цепи.[ …]
Для целлотриозы были выращены очень тонкие монокристаллы, однако провести анализ дальше определения вероятной пространственной группы не удалось [70]. Данные по инфракрасным спектрам и рентгенограмма порошка указывают, что упаковка цепей в целлодекстринах, начиная с целлотетраозы, очень похожа на упаковку цепей в целлюлозе II, хотя период идентичности вдоль оси цепи зависит для низших гомологов от длины молекулы [71]. Анализ монокристаллов целлотетраозы и целлопентаозы может дать очень интересные данные, однако он очень утомителен. Согласно сообщению [41] об изучении двумерной проекции (х, г; размеры осей такие же, как, указаны в табл. 1.13), кристаллы целлотетраозы имеют триклинную элементарную ячейку, длина которой в два раза больше длины ангидроцеллобиозного звена макромолекулы. Эти предварительные данные для монокристалла могут служить подтверждением того, что элементарная ячейка целлюлозы II является триклинной и содержит параллельно ориентированные цепи.[ …]
К ароматическим УВ относятся как собственно ароматические структуры — 6-членные кольца из радикалов -СН-, так и “гибридные” структуры, состоящие из ароматических и нафтеновых колец. Основная масса ароматических структур составляют моноядерные УВ — гомологи бензола. Полициклические ароматические углеводороды (ПАУ) с двумя и более ароматическими кольцами составляют в нефти 1- 4 %. Среди голоядерных ПАУ большое внимание обычно уделяется 3,4-бензо-(а)пирену как наиболее распространенному представителю канцерогенных веществ.[ …]
Основная масса ароматических структур составляют моноядерные УВ — гомологи бензола. Полициклические ароматические углеводороды (ПАУ) с двумя и более ароматическими кольцами составляют в нефти 1- 4 %. Среди голоядерных ПАУ большое внимание обычно уделяется 3,4-бензо-(а)пирену как наиболее распространенному представителю канцерогенных веществ.[ …]
Трубки для определения окиси углерода 145 могут быть применены для количественного определения других соединений углерода 46. Известно, что взвесь I2O5 в олеуме реагирует не только с окисью углерода, но также с ацетиленом, этиленом, бутаном, бензолом, его гомологами и сероуглеродом. В зависимости от концентрации S03 появляется коричневая, оливково-зеленая или голубоватозеленая окраска, по которой можно определить содержание СО. С помощью таких трубок 145 можно также количественно определить ацетилен, этилен, бутан, бензол, его гомологи и сероуглерод. Фильтрующий патрон применять не нужно.[ …]
Высокой газонасыщенностью обладают воды не-окома на Березовской площади (1844 см3/л), в газах содержится 88% метана и 4,33% его гомологов. Увеличивается также газонасыщенность вод от 300 до 1072 см3/л. Доля гомологов метана в рассматриваемых газах достигает 12,5% (площадь Джанкой).[ …]
Увеличивается также газонасыщенность вод от 300 до 1072 см3/л. Доля гомологов метана в рассматриваемых газах достигает 12,5% (площадь Джанкой).[ …]
Воздействие на окружающую среду. Ароматические УВ -наиболее токсичные компоненты нефти. В концентрации всего 1 % в воде они убивают все водные растения. Нефть, содержащая до 35 % ароматических УВ, значительно угнетает рост высших растений. Моноядерные УВ — бензол и его гомологи оказывают более быстрое токсическое воздействие на организмы чем ПАУ, так как ПАУ медленнее проникают через мембраны клеток. Однако, в целом, ПАУ действуют более длительное время, являясь хроническими токсикантами. Ароматические УВ трудно поддаются разрушению. Экспериментально доказано, что главным фактором деградации ПАУ в окружающей среде, в особенности в воде и воздухе, является фотолиз, инициированный ультрафиолетовым излучением. В почве этот процесс может происходить только на ее поверхности.[ …]
Расщепления хромомер и нитей не отмечают, но считают, что хромосомы на уровне этой стадии являются двойными, т. е. диплоидными. Гомологи каждой хромосомной пары объединяются хромомерами вдоль их длины по принципу застежки.[ …]
е. диплоидными. Гомологи каждой хромосомной пары объединяются хромомерами вдоль их длины по принципу застежки.[ …]
Гёте описал пе только постепенное изменение ряда признаков листьев, последовательно образуемых побегом. В своей теории метаморфоза Гёте показал, что цветки представляют собой мета-морфизироиашше вегетативные побеги, которые вместо зеленых лист],ев несут гга себе пестики, тычинки и цветочные покровы. Эта концепция гомологии между вегетативными и репродуктивными побегами широко распространена среди морфологов растений и в паши дни [Берпье и др., 1985]. Действительно, переходные формы между листьями и прицветниками, чашелистиками и лепестками, лепестками и тычинками, тычинками и пестиками являются обычными для многих видов и родов растений, а также встречаются у многочисленных тератологических форм.[ …]
Слабые погоны содержат много воды, легколетучие примеси и уксусную кислоту. Чем больше воды отгоняется со слабыми погонами, тем концентрированнее получаются крепкие погоны, которые представляют собой техническую уксусную кислоту и сырец ледяной уксусной кислоты или пищевой кислоты. В хвостовых погонах содержится много гомологов уксусной кислоты. Кислый смолистый остаток является наиболее тяжелокипящей частью черной уксусной кислоты.[ …]
В хвостовых погонах содержится много гомологов уксусной кислоты. Кислый смолистый остаток является наиболее тяжелокипящей частью черной уксусной кислоты.[ …]
Установленные особенности изменения качественных и количественных характеристик газов подземных вод позволяют считать параметры газоносности вод надежными показателями региональной и локальной оценки перспектив нефтегазоносности. К разряду прямых показателей относят повышенное по сравнению с фоном количество метана и его гомологов, газонасьнценность вод углеводородными газами, общую и парциальную (по углеводородным газам) упругость газов. Наличие гомологов метана, в особенности бутана, пентана, гексана, в газах, кроме того, может указывать и на характер залежи (нефтяная или газовая). Другие газовые показатели — повышенное содержание «безаргонного» азота и диоксида углерода, коэффициенты СН4/ТУ, СН4/С2Н6, СН4/С3Н8, /-С4Н10/л-С4Н10, /-С5Н12/л-С5Н12, N2/yB, 6 3С, Не/Ar являются косвенными.[ …]
Молекулярные сита адсорбируют практически все вещества, молекулы которых меньше или одинаковы с диаметром пор сита. Молекулы с несколько большим диаметром поглощаются частично, и сорбция их протекает значительно медленнее. Относительная сорбция гомологов парафина зависит от их молекулярной массы. Для воды эта величина равна 2,6,т.е. вода сорбируется в первую очередь.[ …]
Молекулы с несколько большим диаметром поглощаются частично, и сорбция их протекает значительно медленнее. Относительная сорбция гомологов парафина зависит от их молекулярной массы. Для воды эта величина равна 2,6,т.е. вода сорбируется в первую очередь.[ …]
Скелет конечностей имеет особенности, причем ключевым является вопрос о переходе плавников в конечности. Конечности классифицируют на непарные и парные. Примером непарных конечностей является спинной и хвостовой плавники у рыб, парных — свободные конечности, роль которых выполняют плавники у рыб и пятипалые конечности у наземных животных. Парные плавники кистеперых рыб содержат гомологи плечевой, локтевой и лучевой костей пятипалой конечности наземных позвоночных, что свидетельствует о происхождении пятипалой конечности от плавника кистеперых рыб. Свободные конечности четвероногих обычно состоят из трех сегментов, которые организованы по-разному у разных животных. Главные вариации отмечаются в строении костей плеча и бедра. В частности, плечевая и бедренная кости укорочены у дельфинов и некоторых ископаемых форм (ихтиозавров). [ …]
[ …]
В Прикумской зоне поднятий Восточного Предкавказья, где выявлено основное количество нефтяных залежей в известняках триаса, распространены застойные воды хлоридно-кальциевого типа с минерализацией от 40 до 157 г/л (чаще 80—100 г/л), rNa/rCI 0,48—0,90. Газонасыщенность вод колеблется от 1420 до 2990 см3/л. В водорастворенных газах содержится метана 40—70%, повышенное количество углекислоты (10—50%), доля гомологов метана 5—15%.[ …]
На стадии пахинемы, которая является наиболее долгой во времени стадией в мейотической профазе, происходит конденсация бивалентов и разделение каждой хроматиды надвое, в результате чего каждый бивалент представляет собой сложную спиральную структуру, состоящую из четырех сестринских хроматид (тетрад). В конце этой стадии начинается разделение спаренных хромосом-бивалентов. Теперь гомологичные хромосомы могут наблюдаться рядом. Поэтому в некоторых препаратах можно видеть четыре хромосомы, которые образуются в результате дупликации каждого гомолога, формирующего сестринские хроматиды. На этой стадии происходят обмены между гомологами и формирование хиазм.[ …]
На этой стадии происходят обмены между гомологами и формирование хиазм.[ …]
При 1-минутном кипячении концентрация Н2504, равная 68,4%, является оптимальной для процесса окисления. В случае окисления без катализатора кривая во всех случаях практически выравнивается. При окислении о-крезола и смеси ксиленолов, с применением катализатора кривая сохраняет тенденцию умеренно повышаться: так, у о-крезола повысится степень окисления при концентрации Н2504, равной 71 % (по отношению к окислению при концентрации 64,8%), на 4,8% и при 78% Нг304 на 9%; у смеси ксиленолов — при 71% Н2504 на 6,1%, а при 78% Н2504 на 11%. Влияние катализатора при 1-минутном кипячении у гомологов фенола проявляется в большей мере, чем при использовании только реакционного тепла, но главным образом эта величина сказьь вается при более высоких концентрациях Н25 04.[ …]
Источником загрязнения атмосферного воздуха также является пыль, которая выделяется при приготовлении буровых растворов из порошкообразных материалов (неорганизованные выбросы). К этой же категории можно отнести выбросы углеводородов, попадающих в циркуляционную систему буровой установки и в шламовые амбары при возможных нефтегазопроявлениях в процессе бурения или углубления (ремонта) скважин. Среди газов, выделяющихся вместе с углеводородами, могут быть меркаптан и окись углерода, обладающие сильнодействующими свойствами, а бенз(а)пирен и его гомологи признаны канцерогенами. Необходимо учитывать высокую миграционную способность газообразных веществ. Максимальный ореол рассеяния (до 15 км) присущ углеводородам, аммиаку и окислам углерода. Сероводород мигрирует до 5-10 км, а окись азота и серный ангидрид — до 1-3 км от очага загрязнения.[ …]
К этой же категории можно отнести выбросы углеводородов, попадающих в циркуляционную систему буровой установки и в шламовые амбары при возможных нефтегазопроявлениях в процессе бурения или углубления (ремонта) скважин. Среди газов, выделяющихся вместе с углеводородами, могут быть меркаптан и окись углерода, обладающие сильнодействующими свойствами, а бенз(а)пирен и его гомологи признаны канцерогенами. Необходимо учитывать высокую миграционную способность газообразных веществ. Максимальный ореол рассеяния (до 15 км) присущ углеводородам, аммиаку и окислам углерода. Сероводород мигрирует до 5-10 км, а окись азота и серный ангидрид — до 1-3 км от очага загрязнения.[ …]
Рассмотрение основных источников природного сырья показывает, что в большинстве случаев кроме необходимого компонента в них содержатся другие ценные вещества. Например, в железной руде часто присутствуют медь, титан, ванадий, кобальт, цинк, фосфор, сера, свинец и другие редкие элементы. В полиметаллических рудах содержится более 50 ценных элементов, в том числе олово, медь, кобальт, вольфрам, молибден, серебро, золото, металлы платиновой группы. Часто сопутствующие элементы обладают большей ценностью, чем основные, ради которых организовано производство. В природном газе содержатся азот, гелий, сера, а в составе газообразного конденсата — гомологи метана. В нефтях имеются различные соединения серы, и им сопутствуют газы, содержащие ценные углеводороды, а также пластовые воды, в которых находятся иод, бром и бор.[ …]
Часто сопутствующие элементы обладают большей ценностью, чем основные, ради которых организовано производство. В природном газе содержатся азот, гелий, сера, а в составе газообразного конденсата — гомологи метана. В нефтях имеются различные соединения серы, и им сопутствуют газы, содержащие ценные углеводороды, а также пластовые воды, в которых находятся иод, бром и бор.[ …]
Меры предупреждения. Основное — герметизация аппаратуры, в которой происходит получение или переработка соединений данной группы. В работе Навроцкого и др. содержатся конкретные предложения о переустройстве аппаратуры в производстве ароматических нитро- и аминосоединений, представляющей наибольшую опасность отравлений. Механизация внутризаводской транспортировки соединений данной группы (см. в работе Штромберга и Игнатьева схему центральных анилиновых и нитробензоловых станций). Перевозку по железной дороге жидких соединений в бочках заменить перевозкой в герметичных цистернах. См. также «Правила о порядке чистки, осмотра и ремонта аппаратов химических производств» (обязательное постановление НКТ РСФСР № 6 от 7 февраля 1933 г. [162]). Вытяжная вентиляция в местах, где при открывании аппаратуры и других операциях возможно выделение паров в воздух рабочего помещения. Об устройстве вентиляции в производстве амино- и нитросоединений см. работу Навроцкого и др.[ …]
[162]). Вытяжная вентиляция в местах, где при открывании аппаратуры и других операциях возможно выделение паров в воздух рабочего помещения. Об устройстве вентиляции в производстве амино- и нитросоединений см. работу Навроцкого и др.[ …]
Получение и свойства рекомбинантного протеолитически активного гомолога сериновой пептидазы SerPh222 Tenebrio molitor
456
ПРИКЛАДНАЯ БИОХИМИЯ И МИКРОБИОЛОГИЯ том 57 № 5 2021
ТЕРЕЩЕНКОВА и др.
показано, что активность SerPh222 выше при
значении рН 7.9.
Таким образом, разработан метод получения
рекомбинантного белка-гомолога сериновых
пептидаз proSerPh222 T. mol itor, подобраны усло-
вия его очистки с использованием металл-хелат-
ной аффинной хроматографии, проведены дегли-
козилирование эндогликозидазой Endo H и про-
цессинг пробелка до зрелой формы. Показано, что:
(1) полученный гомолог подвергался процессингу
под действием трипсина, (2) проявлял невысокую,
но достоверно детектируемую химотрипсин-по-
добную активность при использовании в качестве
субстрата Suc-Ala-Ala-Pro-Phe-pNA, причем при
рН 7. 9, в слабощелочных условиях, характерных
9, в слабощелочных условиях, характерных
для содержимого задней части средней кишки ли-
чинок T. mo l itor, активность SerPh222 значитель-
но выше, чем при рН 5.6, в условиях содержимого
передней части средней кишки, (3) гликозилиро-
вание не оказывало влияния на изученные свой-
ства. Настоящая работа вносит вклад в исследо-
вание малоизученной области функционирова-
ния псевдопептидаз и может послужить основой
для выяснения связи между структурой и функ-
цией сериновых пептидаз и их гомологов.
Авторы выражают благодарность к. х. н. М.В. Се-
ребряковой за проведение масс-спектрометриче-
ского анализа препарата рекомбинантного гомо-
лога proSerPh222.
Исследование выполнено при финансовой
поддержке гранта Российского научного фонда
(проект № 19-74-00080).
СПИСОК ЛИТЕРАТУРЫ
1. Rawlings N.D., Barrett A.J., Thomas P.D., Huang X.,
Bateman A., Finn R.D. // Nucleic Acids Res. 2018.
V. 46. № D1. P. D624–D632.
https://doi.org/10.1093/nar/gkx1134
2. Brown J.R., Hartley B.S. // Biochem. J. 1966. V. 101.
№ 1. P. 214–228.
https://doi.org/10.1042/bj1010214
3. Perona J.J., Hedstrom L., Rutter W.J., Fletterick R.J. //
Biochemistry. 1995. V. 34. № 5. P. 1489–1499.
https://doi.org/10.1021/bi00005a004
4. Perona J.J., Craik C.S. // Protein Sci. 1995. V. 4. № 3.
P. 337–360.
https://doi.org/10.1002/pro.5560040301
5. Wex T., Lipyansky A., Brömme N.C., Wex H., Guan X.Q.,
Brömme D. // Biochemistry. 2001 V. 40. № 5.
P. 1350–1357.
https://doi.org/10.1021/bi002266o
6. Wu C.Y., Huang J.M., Zhao Y.J., Xu Z.W., Zhu J.Y. //
Toxicon. 2020. V. 183. P. 29–35.
https://doi.org/10.1016/j.toxicon.2020.05.011
7. Pils B., Schultz J. // J. Mol. Biol. 2004. V. 340. № 3.
P. 399–404.
https://doi.org/10.1016/j.jmb.2004.04.063
8. Yang L., Lin Z., Fang Q., Wang J., Yan Z. , Zou Z., Song Q.,
, Zou Z., Song Q.,
Ye G. // Dev. Comp. Immunol. 2017. V. 77. P. 56–68.
https://doi.org/10.1016/j.dci.2017.07.014
9. Reynolds S.L., Fischer K. // Biochem. J. 2015. V. 468.
№ 1. P. 17–24.
https://doi.org/10.1042/BJ20141506
10. Bryant B., Ungerer M.C., Liu Q., Waterhouse R.M.,
Clem R.J. // Insect Biochem. Mol. Biol. 2010. V. 40.
№7. P. 516–523.
https://doi.org/10.1016/j.ibmb.2010.04.011
11. Martynov A.G., Elpidina E.N., Perkin L., Oppert B. //
BMC Genomics. 2015. V. 16. № 75.
https://doi.org/10.1186/s12864-015-1306-x
12. Naumov G.I., Naumova E.S., Tyurin O.V., Kozlov D.G. //
Antonie Van Leeuwenhoek. 2013. V. 104. № 3. P. 339–
347.
https://doi.org/10.1007/s10482-013-9956-7
13. Горбунов А.А., Акентьев Ф.И., Губайдуллин И.И.,
Жиганов Н.И., Терещенкова В.Ф., Элпидина Е.Н.,
Козлов Д.Г. // Биотехнология. 2020. Т. 36. № 6.
С. 78–86.
https://doi.org/10.21519/0234-2758-2020-36-6-78-86
14. Матвеева А.Ю., Губайдуллин И.И., Федоров А.С.,
Матвеева А.Ю., Губайдуллин И.И., Федоров А.С.,
Козлов Д.Г. // Биотехнология. 2019. Т. 35. № 5.
С. 3–11.
https://doi.org/10.21519/0234-2758-2019-35-5-3-11
15. Kozlov D.G., Yagudin T.A. // Biotechnol. Lett. 2008.
V. 30. № 9. P. 1661–1663.
https://doi.org/10.1007/s10529-008-9739-9
16. Sievers F., Wilm A., Dineen D., Gibson T.J., Karplus K.,
Li W., Lopez R., McWilliam H., Remmert M., Söding J.,
Thompson J.D., Higgins D.G. // Mol. Syst. Biol. 2011.
V. 7. P. 539.
https://doi.org/10.1038/msb.2011.75
17. Nicholas K.B., Nicholas H.B. Jr., Deerfield D.W. II. //
Embnew. News. 1997. V. 4. P. 14.
http://www.psc.edu/biomed/genedoc
18. Almagro Armenteros J.J., Tsirigos K.D., Sonderby C.K.,
Petersen T.N., Winther O., Brunak S., von Heijne G.,
Nielsen H. // Nat. Biotechnol. 2019. V. 37. № 4.
P. 420–423.
https://doi.org/10.1038/s41587-019-0036-z
19. Терещенкова В.Ф., Клячко Е.В., Беневоленский С. В.,
В.,
Белозерский М.А., Дунаевский Я.Е., Филиппова И.Ю.,
Элпидина Е.Н. // Прикл. биохимия и микробиоло-
гия. 2019. Т. 55. № 3. С. 231–236.
20. Frugoni J.A.C. // Gazz. Chem. Ital. 1957. V. 87.
P. 403–407.
21. Parry M.A., Fernandez-Catalan C., Bergner A., Huber R.,
Hopfner K.P., Schlott B., Gührs K.H., Bode W. // Nat.
Struct. Biol. 1998. V. 5. № 10. P. 917–923.
https://doi.org/10.1038/2359
22. Hedstrom L. //Chem. Rev. 2002. V. 102. № 12.
P. 4501–4524.
https://doi.org/10.1021/cr000033x
23. Vinokurov K.S., Elpidina E.N., Oppert B., Prabhakar S.,
Zhuzhikov D.P., Dunaevsky Y.E., Belozersky M.A. //
Comp. Biochem. Physiol. B. Biochem. Mol. Biol.
2006. V. 145. № 2. P. 126–137.
https://doi.org/10.1016/j.cbpb.2006.05.005
Задачи к разделу изомерия и номенклатура органических соединений
Здесь представлены типовые задачи к разделу изомерия и номенклатура органических соединений.
1.
 Для вещества, имеющего строение CH3 – CH (СН3) – CH3, составьте формулы одного изомера и двух гомологов. Дайте всем веществам названия по систематической номенклатуре.
Для вещества, имеющего строение CH3 – CH (СН3) – CH3, составьте формулы одного изомера и двух гомологов. Дайте всем веществам названия по систематической номенклатуре.Показать решение »
Изомеры – вещества, обладающие одинаковой молекулярной формулой, но имеющие различное строение и, вследствие этого, различные химические и физические свойства.
Гомологи – вещества, принадлежащие одному гомологическому ряду. Имеют схожее однотипное строение, и каждый последующий член гомологического ряда отличается от предыдущего на – СН2 группу.
2. Для вещества, имеющего строение CH
3 – CH(СН3)– CH2 – CH(СН3) – CH3, составьте формулы одного изомера и двух гомологов. Дайте всем веществам названия по систематической номенклатуре.Показать решение »
Изомеры – вещества, обладающие одинаковой молекулярной формулой, но имеющие различное строение и, вследствие этого, различные химические и физические свойства.
Гомологи – вещества, принадлежащие одному гомологическому ряду. Имеют схожее однотипное строение, и каждый последующий член гомологического ряда отличается от предыдущего на – СН2 группу.
3.Выведите формулы альдегидов и кетонов, изомерных валериановому альдегиду С
5Н10О, и назовите их по рациональной и систематической номенклатурам.Показать решение »
4.Напишите проекционные формулы всех возможных стереомеров следующих соединений: 1,2-дибромпропана; 3,4-дибром-3,4-диметилгексана; 2,4-дибромпентана; 2-бром-З-хлорбутана.
Показать решение »
5.Укажите, будет ли сохранена оптическая активность, если втор-бутилкарбинол: а)окислить в соответствующую карбоновую кислоту; б) превратить в эфир уксусной кислоты; в) превратить в йодистый алкил и последний восстановить до углеводорода?
Показать решение »
Среди органических соединений есть оптически активные вещества, обладающие способностью вращать плоскость поляризации света. Такие вещества существуют в виде двух изомеров (оптических антиподов), вращающих плоскость поляризации в противоположные стороны. Пространственные формы оптических антиподов не совпадают и их совмещение невозможно. Оптическая изомерия возможно только для молекул, имеющих асимметрический атом углерода (т.е. атом углерода, связанный с четырьмя различными заместителями).
Такие вещества существуют в виде двух изомеров (оптических антиподов), вращающих плоскость поляризации в противоположные стороны. Пространственные формы оптических антиподов не совпадают и их совмещение невозможно. Оптическая изомерия возможно только для молекул, имеющих асимметрический атом углерода (т.е. атом углерода, связанный с четырьмя различными заместителями).
Все продукты реакций, кроме, 2-метилбутана, будут проявлять оптическую активность, связанную с наличием в молекуле асимметрического атома углерода (*С)
6.Напишите графические формул пяти изомерных аминов, содержащих бензольное ядро, состава C
7H9N. Назовите их.Показать решение »
7.Напишите графические формулы изомерных ароматических кислот состава С
8Н8О2. Назовите эти соединения.Показать решение »
8.Напишите графические формулы первичных, вторичных и третичных спиртов состава C
6H13OH. Назовите их по номенклатуре ИЮПАК.
Назовите их по номенклатуре ИЮПАК.Показать решение »
9.Сколько изомерных гомологов бензола может отвечать формуле C
9H12? Напишите структурные формулы изомеров и назовите их.Показать решение »
Типы гомологии
Используя генные деревья, мы можем вывести следующие парные отношения.
Ортологи
Гомологи, которые разошлись в результате видообразования. Существует четыре типа ортологов:
- Ортологи 1-к-1 (ortholog_one2one)
- Ортологи 1-ко-многим (ortholog_one2many)
- ортологов многие ко многим (ortholog_many2many)
- межвидовых паралогов – только в виде исключения
Гены разных видов, связанные событием видообразования, определяются как ортологов .В зависимости от количества генов, обнаруженных у каждого вида, мы различаем отношения 1:1, 1:многие и многие:многие. Пожалуйста, обратитесь к рисунку, где приведены примеры трех видов.
Гомеологи
Гомологи, которые разошлись в результате видообразования, но оказались в одном и том же геноме в результате гибридизации (например, полиплоидные геномы растений). Они могут иметь те же типы, что и ортологи (см. выше), но, что важно, типы относятся к количеству генов в субгеномах.
Возьмем в качестве примера пшеницу ( Triticum aestivum ). Это гибрид трех субгеномов, названных A, B и D. Если ген обнаруживается в одной копии в каждом субгеноме, все три гомеолога AB, AD и BD помечаются как гомеологи 1:1. Если у гена есть вторая копия в субгеноме D, пара AB помечена как 1:1, а пары AD и B-D помечены как 1:много.
Ортологи против полиплоидных геномов
Ортологи против полиплоидного генома маркируются с учетом количества генов в субгеномах.Например, при сравнении гексаплоидной пшеницы Triticum aestivum с ячменем Hordeum vulgare ген, который находится в одной копии в каждом субгеноме и в ячмене, будет иметь три ортологических пары 1:1 между пшеницей и ячменем. Если в ячмене две копии, то будет 3×2=6 1:много пар ортологий.
Если субгеном A пшеницы также имеет две копии гена, его пары ортологии с ячменем будут многие: многие, тогда как пары ортологии, включающие субгеномы B и D, все равно будут 1: много.
Если в ячмене две копии, то будет 3×2=6 1:много пар ортологий.
Если субгеном A пшеницы также имеет две копии гена, его пары ортологии с ячменем будут многие: многие, тогда как пары ортологии, включающие субгеномы B и D, все равно будут 1: много.
Паралоги
Гомологи, которые разошлись в результате дупликации. Есть два типа паралогов:
- одновидовые паралогии (inin_species_paralog)
- фрагментов одного и того же предполагаемого гена (gene_split)
Гены одного и того же вида, связанные событием дупликации, определяются как паралогов . На предыдущем рисунке Hsap2 и Hsap2′, а также Mmus2 и Mmus2′ являются двумя примерами внутривидовых паралогов. Событие дублирования, связанное с паралогами, не обязательно должно затрагивать только этот вид.Например, Mmus2′ и Mmus3′ также находятся в паралогах видов, но событие дупликации произошло у общего предка между видами Hsap (человек) и видами Mmus (мышь). Уровень таксономии «умножает» событие дублирования на предка Euarchontoglires.
Показатели уверенности
Мы вычисляем показатель достоверности дублирования для каждого узла дублирования. Это индекс Жаккара наборов видов под двумя поддеревьями. Оценка иногда упоминается как «оценка пересечения видов».
Паралоги между видами
Межвидовой паралог соответствует отношению между генами разных видов, где предковый узел был помечен как узел дупликации, т.е. Mmus1:Hsap2 или Mmus1:Hsap3. В настоящее время мы аннотируем паралог между видами только тогда, когда нет лучшего совпадения ни для одного из генов, а дупликация слабо поддерживается (показатель достоверности дупликации ≤ 0,25).
Такие случаи могут быть результатом реальных дупликаций с последующей потерей генов (как показано на рисунке ниже), но чаще всего они возникают в результате неправильной топологии дерева генов с фиктивным узлом дупликации.Часто за этими проблемами стоят ошибки сборки. Неясно, являются ли эти гены настоящими ортологами или нет, но они являются лучшими доступными кандидатами (с учетом данных), и мы искажаем определение ортологии, чтобы пометить их как ортологи.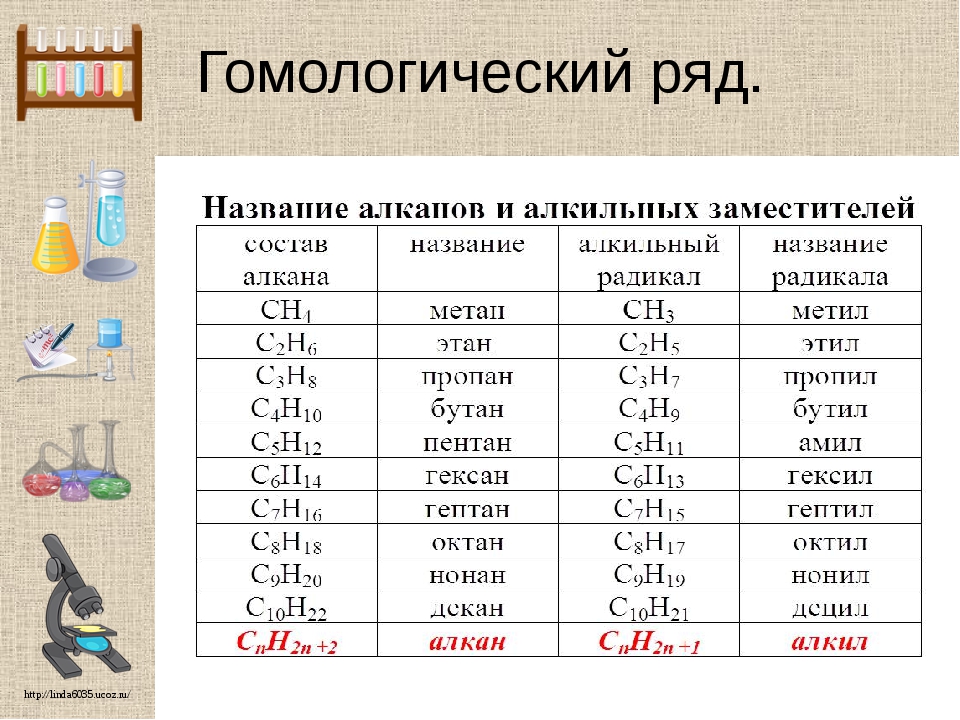 Они помечены как «несовместимые с генным деревом». Людям, интересующимся филогенетическим анализом, смешивающим ортологии и деревья, вероятно, следует использовать набор ортологий, совместимых с деревьями.
Они помечены как «несовместимые с генным деревом». Людям, интересующимся филогенетическим анализом, смешивающим ортологии и деревья, вероятно, следует использовать набор ортологий, совместимых с деревьями.
Ген расщепляется
Паралог, помеченный как расщепление гена, возникает, когда ген оказывается разделенным надвое.В процессе построения генного дерева каждый из них будет выравниваться с одним концом другого ортолога гена, поэтому они будут сгруппированы и размещены в дереве вместе с другими гомологами. Они обычно связаны с фрагментированными сборками генома или предсказанием генов, которое плохо подтверждается доказательствами, что приводит к двум фрагментированным генам там, где должен быть один большой.
Сразу после построения множественного выравнивания мы обнаруживаем пары пар генов, которые лежат близко друг к другу (
Оценка достоверности
Мы проводим ряд дополнительных анализов для оценки качества предсказания ортологии, следуя внешним источникам данных (локальная синтения и выравнивание всего генома). Оттуда мы можем пометить некоторые ортологии как высоконадежные.
Оттуда мы можем пометить некоторые ортологии как высоконадежные.
Функция, гомология и индивидуация признаков* в JSTOR
АбстрактныйЯ защищаю мнение, что многие биологические категории определяются гомологией, против ряда аргументов, направленных на то, чтобы показать, что все биологические категории определяются, по крайней мере частично, выбранной функцией. Я показываю, что категории гомологии «включают аномалии» — нечто, что часто считается уникальным для отдельных категорий функций.Я показываю, что классификации по выбранной функции логически зависят от классификаций по гомологии, но не наоборот. Наконец, я отвергаю точку зрения, что биологи должны использовать соображения избранной функции, чтобы абстрагироваться от вариаций и патологий, чтобы сформировать каноническое описание класса биологических систем.
Информация о журнале Текущие выпуски теперь доступны на веб-сайте Chicago Journals. Прочтите последний выпуск. С момента своего основания в 1934 году «Философия науки» вместе с спонсирующим ее обществом, Ассоциацией философии науки, были посвящены продвижению исследований и свободному обсуждению различных точек зрения в области философии науки.Журнал содержит очерки, дискуссионные статьи и рецензии на книги.
Прочтите последний выпуск. С момента своего основания в 1934 году «Философия науки» вместе с спонсирующим ее обществом, Ассоциацией философии науки, были посвящены продвижению исследований и свободному обсуждению различных точек зрения в области философии науки.Журнал содержит очерки, дискуссионные статьи и рецензии на книги.
С момента своего основания в 1890 году в качестве одного из трех основных подразделений Чикагского университета издательство University of Chicago Press взяло на себя обязательство распространять научные знания самого высокого уровня и публиковать серьезные работы, которые способствуют образованию, способствуют общественному пониманию. и обогатить культурную жизнь. Сегодня Отдел журналов издает более 70 журналов и периодических изданий в твердом переплете по широкому кругу академических дисциплин, включая социальные науки, гуманитарные науки, образование, биологические и медицинские науки, а также физические науки.
Этот предмет является частью коллекции JSTOR.
Условия использования см. в наших Условиях использования
Copyright 2006 Ассоциация философии науки. Все права защищены.
Запросить разрешения
Идентификация гомологов в незначительных бласт-хитах с использованием свойств внешних генов | BMC Биоинформатика
Кунин Е.В. Ортологи, паралоги и эволюционная геномика. Annu Rev Genet 2005, 39: 309–338. 10.1146/annurev.genet.39.073003.114725
CAS Статья пабмед Google Scholar
Копли Р.Р., Гудштадт Л., Понтинг С. Эволюция эукариотического домена на основе сравнения геномов. Curr Opin Genet Dev 2003, 13 (6): 623–628. 10.1016/j.gde.2003.10.004
CAS Статья пабмед Google Scholar

Альтшул С.Ф., Гиш В., Миллер В., Майерс Э.В., Липман Д.Дж.: Базовый инструмент локального поиска центровки. J Mol Biol 1990, 215(3):403–410.
КАС Статья пабмед Google Scholar
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ: Gapped BLAST и PSI-BLAST: новое поколение программ поиска белковых баз данных. Nucleic Acids Res 1997, 25(17):3389–3402. 10.1093/нар/25.17.3389
Центральный пабмед КАС Статья пабмед Google Scholar
Eddy SR: Профиль скрытых марковских моделей. Биоинформатика 1998, 14(9):755–763. 10.1093/биоинформатика/14.9.755
CAS Статья пабмед Google Scholar
Шридхар Дж., Рафи З.А.: Идентификация малых РНК у Enterobacteriaceae с использованием синтении и сохранения геномной основы. Омикс 2007, 11(1):74–99. 10.1089/оми. 2006.0006
2006.0006
CAS Статья пабмед Google Scholar
Татусов Р.Л., Федорова Н.Д., Джексон Дж.Д., Якобс А.Р., Кирютин Б., Кунин Е.В., Крылов Д.М., Мазумдер Р., Мехедов С.Л., Никольская А.Н., Рао Б.С., Смирнов С., Свердлов А.В., Васудеван С., Вольф Ю.И., Инь Ж.Дж., Натале DA: База данных COG: обновленная версия включает эукариоты. BMC Bioinformatics 2003, 4: 41. 10.1186/1471-2105-4-41
PubMed Central Статья пабмед Google Scholar
Курсула П., Оджала Дж., Ламбейр А.М., Виренга Р.К.: Каталитический цикл биосинтетической тиолазы: конформационное путешествие ацетильной группы через четыре способа связывания и две оксианионные дыры.Биохимия 2002, 41(52):15543–15556. 10.1021/би0266232
КАС Статья пабмед Google Scholar
Huynen MA, Snel B, Bork P: Инверсии и динамика эукариотического порядка генов. Тенденции Genet 2001, 17 (6): 304–306. 10.1016/S0168-9525(01)02302-2
Тенденции Genet 2001, 17 (6): 304–306. 10.1016/S0168-9525(01)02302-2
КАС Статья пабмед Google Scholar
Дандекар Т., Снел Б., Хюйнен М., Борк П. Сохранение порядка генов: отпечатки белков, которые физически взаимодействуют.Trends Biochem Sci 1998, 23(9):324–328. 10.1016/С0968-0004(98)01274-2
КАС Статья пабмед Google Scholar
Овербик Р., Фонстейн М., Д’Суза М., Пуш Г.Д., Мальцев Н.: Использование кластеров генов для определения функциональной связи. Proc Natl Acad Sci U S A 1999, 96(6):2896–2901. 10.1073/pnas.96.6.2896
PubMed Central КАС Статья пабмед Google Scholar
Коин Л., Бейтман А., Дурбин Р.: Расширенное обнаружение белковых доменов с использованием таксономии. BMC Bioinformatics 2004, 5: 56. 10.1186/1471-2105-5-56
PubMed Central Статья пабмед Google Scholar

Садреев Р., Гришин Н.: КОМПАС: инструмент для сравнения множественных выравниваний белков с оценкой статистической значимости. J Mol Biol 2003, 326(1):317–336. 10.1016/S0022-2836(02)01371-2
КАС Статья пабмед Google Scholar
Soding J: Определение гомологии белков путем сравнения HMM-HMM. Биоинформатика 2005, 21(7):951–960. 10.1093/биоинформатика/bti125
Статья пабмед Google Scholar
Коин Л., Бейтман А., Дурбин Р.: Усовершенствованное обнаружение белковых доменов с использованием методов языкового моделирования на основе распознавания речи. Proc Natl Acad Sci U S A 2003, 100(8):4516–4520. 10.1073/pnas.0737502100
PubMed Central КАС Статья пабмед Google Scholar
Griffiths E, Ventresca MS, Gupta RS: Скрининг BLAST геномов хламидий для идентификации сигнатурных белков, уникальных для групп видов Chlamydiales, Chlamydiaceae, Chlamydophila и Chlamydia. BMC Genomics 2006, 7: 14. 10.1186/1471-2164-7-14
BMC Genomics 2006, 7: 14. 10.1186/1471-2164-7-14
PubMed Central Статья пабмед Google Scholar
Хартман Х, Федоров А: Происхождение эукариотической клетки: геномное исследование. Proc Natl Acad Sci U S A 2002, 99(3):1420–1425.10.1073/pnas.032658599
PubMed Central КАС Статья пабмед Google Scholar
Макарова К., Слесарев А., Вольф Ю., Сорокин А., Миркин Б., Кунин Е., Павлов А., Павлова Н., Карамычев В., Полушин Н., Шахова В., Григорьев И., Лу Ю., Рохсар Д., Лукас С. , Хуанг К., Гудштейн Д.М., Хокинс Т., Пленгвидья В., Велкер Д., Хьюз Дж., Го Ю., Бенсон А., Болдуин К., Ли Дж. Х., Диас-Мунис И., Дости Б., Смеянов В., Вехтер В., Баработе Р., Лорка Г. , Альтерманн Э., Баррангу Р., Ганесан Б., Се И., Роусторн Х., Тамир Д., Паркер С., Брейдт Ф., Бродбент Дж., Хаткинс Р., О’Салливан Д., Стил Дж., Унлу Г., Сайер М., Клаенхаммер Т., Ричардсон П. , Козявкин С., Веймер Б., Миллс Д.: Сравнительная геномика молочнокислых бактерий.Proc Natl Acad Sci U S A 2006, 103(42):15611–15616. 10.1073/pnas.0607117103
, Козявкин С., Веймер Б., Миллс Д.: Сравнительная геномика молочнокислых бактерий.Proc Natl Acad Sci U S A 2006, 103(42):15611–15616. 10.1073/pnas.0607117103
PubMed Central Статья пабмед Google Scholar
Финн Р.Д., Мистри Дж., Шустер-Боклер Б., Гриффитс-Джонс С., Холлих В., Лассманн Т., Моксон С., Маршалл М., Ханна А., Дурбин Р., Эдди С.Р., Соннхаммер Э.Л., Бейтман А.: Pfam : кланы, веб-инструменты и сервисы. Nucleic Acids Res 2006, 34 (выпуск базы данных): D247–51. 10.1093/nar/gkj149
Центральный пабмед КАС Статья пабмед Google Scholar
Мурзин А.Г., Бреннер С.Е., Хаббард Т., Чотия С.: SCOP: база данных структурной классификации белков для исследования последовательностей и структур. Дж. Мол. Биол. 1995, 247(4):536–540. 10.1006/джмби.1995.0159
КАС пабмед Google Scholar

Гарднер М.Дж., Холл Н., Фунг Э., Уайт О., Берриман М., Хайман Р.В., Карлтон Дж.М., Пейн А., Нельсон К.Е., Боуман С., Полсен И.Т., Джеймс К., Эйзен Дж.А., Резерфорд К., Зальцберг С.Л. , Крейг А., Кайес С., Чан М.С., Нене В., Шаллом С.Дж., Сух Б., Петерсон Дж., Ангиуоли С., Пертеа М., Аллен Дж., Селенгут Дж., Хафт Д., Мазер М.В., Вайдья А.Б., Мартин Д.М., Фэрламб А.Х., Фраунхольц MJ, Roos DS, Ralph SA, McFadden GI, Cummings LM, Subramanian GM, Mungall C, Venter JC, Carucci DJ, Hoffman SL, Newbold C, Davis RW, Fraser CM, Barrell B: Геномная последовательность человеческого малярийного паразита Plasmodium falciparum .Природа 2002, 419 (6906): 498–511. 10.1038/nature01097
CAS Статья пабмед Google Scholar
Кунин Е.В.: Кунин Евгений В. (Интервью). Curr Biol 2004, 14(3):R96–7. 10.1016/S0960-9822(04)00025-9
КАС Статья пабмед Google Scholar

Benson DA, Karsch-Mizrachi I, Lipman DJ, Ostell J, Wheeler DL: GenBank. Nucleic Acids Res 2006, 34 (выпуск базы данных): D16–20.10.1093/nar/gkj157
Центральный пабмед КАС Статья пабмед Google Scholar
Edgar RC: MUSCLE: множественное выравнивание последовательностей с высокой точностью и высокой производительностью. Nucleic Acids Res 2004, 32(5):1792–1797. 10.1093/нар/гх440
Центральный пабмед КАС Статья пабмед Google Scholar
Guindon S, Gascuel O: Простой, быстрый и точный алгоритм для оценки больших филогений по максимальному правдоподобию.Сист Биол 2003, 52(5):696–704. 10.1080/106351503
Артикул пабмед Google Scholar
Page RD: TreeView: приложение для отображения филогенетических деревьев на персональных компьютерах. Comput Appl Biosci 1996, 12(4):357–358.
КАС пабмед Google Scholar
гомология | эволюция | Британика
гомология , в биологии сходство строения, физиологии или развития различных видов организмов, основанное на их происхождении от общего эволюционного предка.Гомология противопоставляется аналогии, которая представляет собой функциональное сходство структуры, основанное не на общем эволюционном происхождении, а просто на сходстве использования. Таким образом, передние конечности таких сильно различающихся млекопитающих, как люди, летучие мыши и олени, гомологичны; форма строения и количество костей в этих различных конечностях практически идентичны и представляют собой адаптивные модификации строения передних конечностей их общих ранних предков-млекопитающих. Аналогичные структуры, с другой стороны, могут быть представлены крыльями птиц и насекомых; структуры используются для полета у обоих типов организмов, но не имеют общего предкового происхождения в начале своего эволюционного развития. Британский биолог XIX века сэр Ричард Оуэн был первым, кто точно определил и гомологию, и аналогию.
Британский биолог XIX века сэр Ричард Оуэн был первым, кто точно определил и гомологию, и аналогию.
Когда два или более органов или структур в основном сходны друг с другом по строению, но модифицированы для выполнения различных функций, говорят, что они серийно гомологичны. Примером этого является крыло летучей мыши и плавник кита. Оба произошли из передних конечностей ранних предков млекопитающих, но они претерпели различные эволюционные модификации для выполнения совершенно разных задач полета и плавания соответственно.Иногда неясно, являются ли сходства в строении у разных организмов аналогичными или гомологичными. Примером этого являются крылья летучих мышей и птиц. Эти структуры гомологичны в том смысле, что в обоих случаях они являются модификациями костной структуры передних конечностей ранних рептилий. Но крылья птиц отличаются от крыльев летучих мышей количеством пальцев и наличием перьев для полета, в то время как у летучих мышей их нет. И самое главное, способность к полету возникла независимо у этих двух различных классов позвоночных; у птиц, когда они эволюционировали из ранних рептилий, и у летучих мышей после того, как их млекопитающие предки уже полностью дифференцировались от рептилий. Таким образом, крылья летучих мышей и птиц можно рассматривать как аналогичные, а не гомологичные, при более тщательном изучении их морфологических различий и эволюционного происхождения.
Таким образом, крылья летучих мышей и птиц можно рассматривать как аналогичные, а не гомологичные, при более тщательном изучении их морфологических различий и эволюционного происхождения.
Организмы, эволюционировавшие разными путями, могут иметь аналогичную структуру, т. е. анатомические особенности, внешне похожие друг на друга (например, крылья птиц и насекомых). Хотя такие структуры выполняют схожие функции, они имеют совершенно разное эволюционное происхождение и модели развития.
Британская энциклопедия, Inc.Подробнее по этой теме
Морфология: Гомология и аналогия
Гомологичные структуры развиваются из сходных эмбриональных субстанций и, таким образом, имеют сходные базовые структуры и модели развития, отражающие…
Эта статья была недавно пересмотрена и обновлена Джоном П. Рафферти.7.13C: гомологи, ортологи и паралоги
Цели обучения
- Различают гомологи, ортологи и паралоги
Гомология составляет основу организации сравнительной биологии. Гомологичный признак часто называют гомологом (также пишется как гомолог). В генетике термин «гомолог» используется как для обозначения гомологичного белка, так и для гена (последовательности ДНК), кодирующего его. Как и в случае с анатомическими структурами, гомология между последовательностями белка или ДНК определяется с точки зрения общего происхождения. Два сегмента ДНК могут иметь общее происхождение либо из-за события видообразования (ортологи), либо из-за события дупликации (паралоги). О гомологии между белками или ДНК часто делается неправильный вывод на основании сходства последовательностей.Термины «процентная гомология» и «сходство последовательностей» часто используются взаимозаменяемо. Как и в случае с анатомическими структурами, высокое сходство последовательностей может возникать из-за конвергентной эволюции или, как в случае с более короткими последовательностями, из-за случайности. Такие последовательности сходны, но не гомологичны. Участки последовательности, которые являются гомологичными, также называются консервативными.
Гомологичный признак часто называют гомологом (также пишется как гомолог). В генетике термин «гомолог» используется как для обозначения гомологичного белка, так и для гена (последовательности ДНК), кодирующего его. Как и в случае с анатомическими структурами, гомология между последовательностями белка или ДНК определяется с точки зрения общего происхождения. Два сегмента ДНК могут иметь общее происхождение либо из-за события видообразования (ортологи), либо из-за события дупликации (паралоги). О гомологии между белками или ДНК часто делается неправильный вывод на основании сходства последовательностей.Термины «процентная гомология» и «сходство последовательностей» часто используются взаимозаменяемо. Как и в случае с анатомическими структурами, высокое сходство последовательностей может возникать из-за конвергентной эволюции или, как в случае с более короткими последовательностями, из-за случайности. Такие последовательности сходны, но не гомологичны. Участки последовательности, которые являются гомологичными, также называются консервативными. Это не следует путать с консервативностью аминокислотных последовательностей, в которой аминокислота в определенном положении заменена другой аминокислотой с функционально эквивалентными физико-химическими свойствами.Однако можно сослаться на частичную гомологию, когда часть сравниваемых последовательностей (предполагается, что они имеют) имеют общее происхождение, а остальные — нет. Например, частичная гомология может быть результатом слияния генов.
Это не следует путать с консервативностью аминокислотных последовательностей, в которой аминокислота в определенном положении заменена другой аминокислотой с функционально эквивалентными физико-химическими свойствами.Однако можно сослаться на частичную гомологию, когда часть сравниваемых последовательностей (предполагается, что они имеют) имеют общее происхождение, а остальные — нет. Например, частичная гомология может быть результатом слияния генов.
Гомологичные последовательности являются ортологичными, если они были разделены событием видообразования: когда вид расходится на два отдельных вида, говорят, что копии одного гена в двух получившихся видах являются ортологичными.Ортологи, или ортологичные гены, — это гены разных видов, возникшие в результате вертикального происхождения от одного гена последнего общего предка. Например, регуляторный белок растительного гриппа присутствует как в Arabidopsis (многоклеточные высшие растения), так и в Chlamydomonas (одноклеточные зеленые водоросли). Вариант Chlamydomonas более сложен: он пересекает мембрану дважды, а не один раз, содержит дополнительные домены и подвергается альтернативному сплайсингу. Однако он может полностью заменить гораздо более простой белок Arabidopsis , если перенести его из водорослей в геном растения с помощью генной инженерии.Значительное сходство последовательностей и общие функциональные домены указывают на то, что эти два гена являются ортологичными генами, унаследованными от общего предка. Ортологичные последовательности предоставляют полезную информацию для таксономической классификации и филогенетических исследований организмов. Модель генетической дивергенции можно использовать для отслеживания родства организмов. Два организма, которые очень тесно связаны между собой, вероятно, будут иметь очень похожие последовательности ДНК между двумя ортологами.
Например, регуляторный белок растительного гриппа присутствует как в Arabidopsis (многоклеточные высшие растения), так и в Chlamydomonas (одноклеточные зеленые водоросли). Вариант Chlamydomonas более сложен: он пересекает мембрану дважды, а не один раз, содержит дополнительные домены и подвергается альтернативному сплайсингу. Однако он может полностью заменить гораздо более простой белок Arabidopsis , если перенести его из водорослей в геном растения с помощью генной инженерии.Значительное сходство последовательностей и общие функциональные домены указывают на то, что эти два гена являются ортологичными генами, унаследованными от общего предка. Ортологичные последовательности предоставляют полезную информацию для таксономической классификации и филогенетических исследований организмов. Модель генетической дивергенции можно использовать для отслеживания родства организмов. Два организма, которые очень тесно связаны между собой, вероятно, будут иметь очень похожие последовательности ДНК между двумя ортологами. И наоборот, организм, эволюционно удаленный от другого организма, скорее всего, будет демонстрировать большее расхождение в последовательности изучаемых ортологов.
И наоборот, организм, эволюционно удаленный от другого организма, скорее всего, будет демонстрировать большее расхождение в последовательности изучаемых ортологов.
Гомологичные последовательности являются паралогичными, если они были разделены событием дупликации гена: если ген в организме дублируется, чтобы занять две разные позиции в одном и том же геноме, то две копии являются паралогичными. Паралогичные гены часто принадлежат одному и тому же виду, но это необязательно. Например, ген гемоглобина человека и ген миоглобина шимпанзе являются паралогами. Паралоги можно разделить на ин-паралоги (паралогические пары, возникшие после видообразования) и внепаралоги (паралогические пары, возникшие до видообразования).Межвидовые аут-паралоги — это пары паралогов, которые существуют между двумя организмами из-за дупликации до видообразования. Внутривидовые внепаралоги — это пары паралогов, которые существуют в одном и том же организме, но дупликация которых произошла после видообразования. Паралоги обычно имеют ту же или аналогичную функцию, но иногда и нет. Из-за отсутствия исходного селективного давления на одну копию дуплицированного гена эта копия может свободно мутировать и приобретать новые функции. Паралогичные последовательности дают полезную информацию о том, как эволюционируют геномы.Гены, кодирующие миоглобин и гемоглобин, считаются древними паралогами. Точно так же четыре известных класса гемоглобинов (гемоглобин А, гемоглобин А2, гемоглобин В и гемоглобин F) являются паралогами друг друга. Хотя каждый из этих белков выполняет одну и ту же основную функцию транспорта кислорода, их функции уже немного различаются: гемоглобин плода (гемоглобин F) имеет более высокое сродство к кислороду, чем гемоглобин взрослого человека. Однако функция не всегда сохраняется. Например, человеческий ангиогенин отделился от рибонуклеазы, и, хотя эти два паралога остаются сходными по третичной структуре, их функции внутри клетки теперь совершенно различны.
Паралоги обычно имеют ту же или аналогичную функцию, но иногда и нет. Из-за отсутствия исходного селективного давления на одну копию дуплицированного гена эта копия может свободно мутировать и приобретать новые функции. Паралогичные последовательности дают полезную информацию о том, как эволюционируют геномы.Гены, кодирующие миоглобин и гемоглобин, считаются древними паралогами. Точно так же четыре известных класса гемоглобинов (гемоглобин А, гемоглобин А2, гемоглобин В и гемоглобин F) являются паралогами друг друга. Хотя каждый из этих белков выполняет одну и ту же основную функцию транспорта кислорода, их функции уже немного различаются: гемоглобин плода (гемоглобин F) имеет более высокое сродство к кислороду, чем гемоглобин взрослого человека. Однако функция не всегда сохраняется. Например, человеческий ангиогенин отделился от рибонуклеазы, и, хотя эти два паралога остаются сходными по третичной структуре, их функции внутри клетки теперь совершенно различны.
Ключевые моменты
- Гомологичный ген (или гомолог) — это ген, унаследованный у двух видов от общего предка. Хотя гомологичные гены могут быть сходными по последовательности, сходные последовательности не обязательно гомологичны.
- Ортологичные — это гомологичные гены, в которых ген расходится после видообразования, но ген и его основная функция сохраняются.
- Если ген дублируется у вида, полученные в результате дублированные гены являются паралогами друг друга, даже если со временем они могут измениться по составу последовательности и функции.
 Хотя гомологичные гены могут быть сходными по последовательности, сходные последовательности не обязательно гомологичны.
Хотя гомологичные гены могут быть сходными по последовательности, сходные последовательности не обязательно гомологичны.Основные термины
- консервативные : В биологии консервативные последовательности — это сходные или идентичные последовательности, которые встречаются в последовательностях нуклеиновых кислот (таких как последовательности РНК и ДНК), белковых последовательностях, белковых структурах.
- селективное давление : Любая причина, снижающая репродуктивный успех в части популяции, потенциально оказывает эволюционное давление или давление отбора.
гомолог — Викисловарь
Английский[править]
Альтернативные формы[править]
Этимология
Заимствован от французского гомолога .
Произношение[править]
Существительное[править]
гомолог ( множественное число гомологи )
- Нечто гомологичное; гомологичный орган или часть, химическое соединение, хромосома, ген или элемент культуры.
- (лингвистика) Слово, используемое в двух языках или диалектах.
- (генетика) Одна из группы похожих последовательностей ДНК, имеющих общее происхождение.
- (органическая химия) Член гомологического ряда.
- (культурная антропология) Убеждение, практика, концепция или артефакт, имеющие аналог в другой культуре.
Связанные термины[править]
Переводы[править]
- Приведенные ниже переводы необходимо проверить и вставить выше в соответствующие таблицы переводов, удалив все номера. Числа не обязательно совпадают с числами в определениях. Инструкции см. в Викисловаре: Макет записи § Переводы.
Переводы для проверки
Этимология
Выученное заимствование из древнегреческого ὁμόλογος (homólogos, «согласие, единомыслие»), эквивалентное homo- + -logue .
Произношение[править]
Прилагательное[править]
гомолог ( множественное число гомологи )
- гомологичный (все значения)
Похожие треугольники, коты гомологи другие пропорции. ― В подобных треугольниках пропорциональны гомологичных сторон.
Существительное[править]
гомолог m или f ( множественное число гомологов )
- аналог
Le ministre s’est entretenu avec son гомолог français. ― Министр провел дискуссию со своим французским коллегой .
Глагол[править]
гомолог
- склонение гомолога («официально признавать, ратифицировать»):
- первое/третье лицо единственного числа настоящее изъявительное/сослагательное наклонение
- повелительное наклонение второго лица единственного числа
Дополнительная литература[править]
Португальский[править]
Глагол[править]
гомолог
- перегиб гомолога («омологировать»):
- первое/третье лицо единственного числа настоящее сослагательное наклонение
- повелительное наклонение от первого/третьего лица единственного числа
испанский язык[править]
Глагол[править]
гомолог
- Формальное второе лицо единственного числа ( usted ) повелительная форма омологара .
- Первое лицо единственного числа ( yo ) форма настоящего сослагательного наклонения от гомолога .
- Формальное второе лицо единственного числа ( usted ) настоящая сослагательная форма омологара .
- Третье лицо единственного числа ( él , ella , также используется с usted ? ) настоящая сослагательная форма гомологара .

В поисках правильного партнера: мейотическое ухаживание
Гомологичные хромосомы обычно разделяются в начале мейоза;
как они образуют пары — одна из выдающихся загадок мейотического
процесс.Уменьшение расстояния между гомологами делает возможным
возникновение хромосомных взаимодействий, ведущих к обнаружению гомологии и
образование бивалентов. У многих организмов хромосома, управляемая теломерами,
генерируются движения, которые сближают гомологи. Дополнительный
движения, вызванные конформационными изменениями хроматина в начале мейоза
также может способствовать гомологичным контактам. Организмы, используемые для изучения
мейоз демонстрирует удивительное разнообразие стратегий обнаружения гомологии.В
двукрылых гомологичные хромосомы остаются парными на протяжении большей части
разработка. Пейринг, по-видимому, возникает как баланс между промоутером и
спаривание генов-супрессоров. Некоторые грибы, растения и животные используют механизмы
на основе рекомбинационных взаимодействий. Другие механизмы, ведущие к гомологии
search не зависят от рекомбинации и требуют специализированных сайтов для спаривания. В
у червя Caenorhabditis elegans каждая хромосома несет центр спаривания
состоящий из хромосомоспецифического ДНК-белкового комплекса, а в делении
дрожжи Schizosaccharomyces pombe , локус sme2 кодирует специфичный для мейоза
некодирующая РНК, которая опосредует гомологичное распознавание.Кроме того,
коррекция несоответствия играет важную роль, особенно у полиплоидов, которые
развились генетические системы, которые подавляют спаривание между негомологичными
родственные (гомеологичные) хромосомы.
Организмы, используемые для изучения
мейоз демонстрирует удивительное разнообразие стратегий обнаружения гомологии.В
двукрылых гомологичные хромосомы остаются парными на протяжении большей части
разработка. Пейринг, по-видимому, возникает как баланс между промоутером и
спаривание генов-супрессоров. Некоторые грибы, растения и животные используют механизмы
на основе рекомбинационных взаимодействий. Другие механизмы, ведущие к гомологии
search не зависят от рекомбинации и требуют специализированных сайтов для спаривания. В
у червя Caenorhabditis elegans каждая хромосома несет центр спаривания
состоящий из хромосомоспецифического ДНК-белкового комплекса, а в делении
дрожжи Schizosaccharomyces pombe , локус sme2 кодирует специфичный для мейоза
некодирующая РНК, которая опосредует гомологичное распознавание.Кроме того,
коррекция несоответствия играет важную роль, особенно у полиплоидов, которые
развились генетические системы, которые подавляют спаривание между негомологичными
родственные (гомеологичные) хромосомы.
1. Введение
Мейоз является центральным процессом в жизненном цикле всех организмов, размножающихся половым путем, который эволюционировал, чтобы компенсировать удвоение числа хромосом, возникающее при оплодотворении, и создавать новые комбинации между родительскими аллелями, повышающие генетическое разнообразие.Зародышевые клетки вступают в мейоз, реплицировав свои хромосомы, и, выполняя два последовательных клеточных деления без промежуточной репликации ДНК, производят гаплоидные гаметы. Зародышевые клетки диплоидных организмов инициируют мейоз, содержащий два набора хромосом, по одному унаследованному от каждого родителя. Гомологичные хромосомы разделяются в первом делении (редукционное деление), а сестринские хроматиды расходятся друг от друга во втором делении (эквационное деление). Для правильной сегрегации хромосом гомологи взаимодействуют и создают устойчивые ассоциации во время профазы I, которые поддерживают их связанными в бивалентной конфигурации до метафазы I. Цитологические структуры, связывающие каждую гомологичную пару в метафазе I, называются хиазмами. Сплоченность сестринских хроматид взаимодействует с хиазмами, обеспечивая стабильность связей между каждой гомологичной парой через метафазу I. Разрушение ядерной оболочки позволяет микротрубочкам взаимодействовать с сестринскими кинетокорами, которые ориентируются на один и тот же полюс веретена. Растворение сплоченности сестринских хроматид, за исключением прицентромерной области, обеспечивает разрешение хиазмы и сегрегацию хромосом в анафазе I.
Цитологические структуры, связывающие каждую гомологичную пару в метафазе I, называются хиазмами. Сплоченность сестринских хроматид взаимодействует с хиазмами, обеспечивая стабильность связей между каждой гомологичной парой через метафазу I. Разрушение ядерной оболочки позволяет микротрубочкам взаимодействовать с сестринскими кинетокорами, которые ориентируются на один и тот же полюс веретена. Растворение сплоченности сестринских хроматид, за исключением прицентромерной области, обеспечивает разрешение хиазмы и сегрегацию хромосом в анафазе I.
Хиазмы образуются после кульминации трех основных процессов, инициированных в ранней профазе I: гомологичное спаривание (т. е. взаимодействие хромосом, приводящее к близкому сближению гомологов по всей их длине), синапсис (т. е. образование белковой синаптонемы). сложная структура между каждой гомологичной парой) и кроссинговер (т.е. реципрокный обмен генетическим материалом между гомологичными хроматидами). Кроссовер и некроссовер (невзаимный обмен) представляют собой два возможных результата, которым следует механизм гомологичной рекомбинации для восстановления одного двухцепочечного разрыва ДНК (DSB), образованного при инициации мейоза. Кроссоверы включают реципрокный обмен последовательностями, фланкирующими сайты репарации, и некроссоверы (также называемые конверсиями генов), перенос локальной информации, охватывающей несколько сотен пар оснований, от одного гомолога к другому. Большинству DSB суждено стать некроссоверными продуктами. Основные этапы механизма рекомбинации обнаружены у дрожжей и консервативны у других эукариот [1]. Мейотическая рекомбинация инициируется консервативным топоизомеразоподобным ферментом Spo11, который вводит запрограммированные DSB в геном.Spo11 разрезает ДНК с помощью реакции, подобной топоизомеразе, для образования ковалентных связей белок-ДНК с 5′-концами ДНК по обе стороны от разрыва. Ферментативный механизм, который восстанавливает DSB, организован таким образом, что предпочтительными в качестве матрицы последовательностями ДНК являются последовательности гомологичной хромосомы, в то время как участие сестринской хроматиды ингибируется. После удаления Spo11 концы DSB затем подвергаются нуклеолитической резекции 5′-цепей с образованием 3′-одноцепочечных хвостов.
Кроссоверы включают реципрокный обмен последовательностями, фланкирующими сайты репарации, и некроссоверы (также называемые конверсиями генов), перенос локальной информации, охватывающей несколько сотен пар оснований, от одного гомолога к другому. Большинству DSB суждено стать некроссоверными продуктами. Основные этапы механизма рекомбинации обнаружены у дрожжей и консервативны у других эукариот [1]. Мейотическая рекомбинация инициируется консервативным топоизомеразоподобным ферментом Spo11, который вводит запрограммированные DSB в геном.Spo11 разрезает ДНК с помощью реакции, подобной топоизомеразе, для образования ковалентных связей белок-ДНК с 5′-концами ДНК по обе стороны от разрыва. Ферментативный механизм, который восстанавливает DSB, организован таким образом, что предпочтительными в качестве матрицы последовательностями ДНК являются последовательности гомологичной хромосомы, в то время как участие сестринской хроматиды ингибируется. После удаления Spo11 концы DSB затем подвергаются нуклеолитической резекции 5′-цепей с образованием 3′-одноцепочечных хвостов. Затем один из этих хвостов может проникнуть в интактную двухцепочечную ДНК гомологичной хромосомы.У большинства эукариот эта реакция, известная как инвазия цепей, требует действия двух рекомбиназ, Rad51 и Dmc1, которые действуют в ассоциации с другими белками [2]. Первоначальные промежуточные соединения вторжения в нити могут быть дополнительно обработаны различными способами с различными результатами рекомбинации продуктов (рис. 1). Если единственный конец DSB, который вторгся в гомологичный партнер, после праймирования синтеза ДНК, смещается и отжигается с другим концом DSB, возникает некроссовер. Этот процесс называется зависимым от синтеза отжигом цепей (SDSA).Альтернативный путь приводит к стабилизации промежуточных звеньев инвазии цепи и захвату второго конца DSB, который инициирует синтез ДНК. При лигировании образуется промежуточное соединение молекулы соединения с двойным соединением Холлидея, которое расщепляется с образованием перекрестного продукта. Кроссоверные и некроссоверные рекомбинационные взаимодействия расходятся при переходе лептотен-зиготена до образования обширных интермедиатов обмена цепями [3, 4].
Затем один из этих хвостов может проникнуть в интактную двухцепочечную ДНК гомологичной хромосомы.У большинства эукариот эта реакция, известная как инвазия цепей, требует действия двух рекомбиназ, Rad51 и Dmc1, которые действуют в ассоциации с другими белками [2]. Первоначальные промежуточные соединения вторжения в нити могут быть дополнительно обработаны различными способами с различными результатами рекомбинации продуктов (рис. 1). Если единственный конец DSB, который вторгся в гомологичный партнер, после праймирования синтеза ДНК, смещается и отжигается с другим концом DSB, возникает некроссовер. Этот процесс называется зависимым от синтеза отжигом цепей (SDSA).Альтернативный путь приводит к стабилизации промежуточных звеньев инвазии цепи и захвату второго конца DSB, который инициирует синтез ДНК. При лигировании образуется промежуточное соединение молекулы соединения с двойным соединением Холлидея, которое расщепляется с образованием перекрестного продукта. Кроссоверные и некроссоверные рекомбинационные взаимодействия расходятся при переходе лептотен-зиготена до образования обширных интермедиатов обмена цепями [3, 4]. Белок Sgs1 почкующихся дрожжей Saccharomyces cerevisiae , гомолог геликазы BLM млекопитающих, который поддерживает стабильность генома, предотвращая накопление аберрантных интермедиатов рекомбинации, является центральным регулятором выбора пути рекомбинации в нормальном мейозе [5, 6].Sgs1 контролирует мейотическую рекомбинацию, предотвращая накопление нерегулируемых интермедиатов суставных молекул. Sgs1 смещает вторгшуюся нить суставных молекул с образованием некроссоверов и предотвращает накопление мультихроматидных суставных молекул. Кроме того, Sgs1 направляет некоторые др. суставные молекулы для взаимодействия с группой специфичных для мейоза белков, называемых семейством ZMM, которые необходимы для образования стабильных суставных молекул и промежуточных соединений двойных соединений Холидея. Эти ZMM-защищенные промежуточные продукты рекомбинации позже, в конце пахитены, разрешаются как кроссинговеры мультибелковым фактором, содержащим нуклеазу Exo1 и комплекс MutL γ Mlh2-Mlh4, который активируется полоподобной киназой Cdc5.
Белок Sgs1 почкующихся дрожжей Saccharomyces cerevisiae , гомолог геликазы BLM млекопитающих, который поддерживает стабильность генома, предотвращая накопление аберрантных интермедиатов рекомбинации, является центральным регулятором выбора пути рекомбинации в нормальном мейозе [5, 6].Sgs1 контролирует мейотическую рекомбинацию, предотвращая накопление нерегулируемых интермедиатов суставных молекул. Sgs1 смещает вторгшуюся нить суставных молекул с образованием некроссоверов и предотвращает накопление мультихроматидных суставных молекул. Кроме того, Sgs1 направляет некоторые др. суставные молекулы для взаимодействия с группой специфичных для мейоза белков, называемых семейством ZMM, которые необходимы для образования стабильных суставных молекул и промежуточных соединений двойных соединений Холидея. Эти ZMM-защищенные промежуточные продукты рекомбинации позже, в конце пахитены, разрешаются как кроссинговеры мультибелковым фактором, содержащим нуклеазу Exo1 и комплекс MutL γ Mlh2-Mlh4, который активируется полоподобной киназой Cdc5. Совместные молекулы, полученные в результате одноцепочечной инвазии, которые ускользают от контроля Sgs1 или продуцируются в его отсутствие, часто образуют мультихроматидные интермедиаты, которые дают как перекрестные, так и неперекрестные продукты, в основном под действием Mus81-Mms4 (MUS81-EME1 у человека). ) комплекс резольвазы, который также нуждается в Cdc5 и в незначительной степени под действием Yen1 (GEN1) или Slx1-Slx4 (BTBD12/SLX4). Другие белки, участвующие в перекрестных и неперекрестных путях, описаны в обзоре [7].
Совместные молекулы, полученные в результате одноцепочечной инвазии, которые ускользают от контроля Sgs1 или продуцируются в его отсутствие, часто образуют мультихроматидные интермедиаты, которые дают как перекрестные, так и неперекрестные продукты, в основном под действием Mus81-Mms4 (MUS81-EME1 у человека). ) комплекс резольвазы, который также нуждается в Cdc5 и в незначительной степени под действием Yen1 (GEN1) или Slx1-Slx4 (BTBD12/SLX4). Другие белки, участвующие в перекрестных и неперекрестных путях, описаны в обзоре [7].
Синаптонемный комплекс представляет собой белковую матрицу, которая в норме образуется между каждой гомологичной парой, усиливая их взаимодействие, и встречается у большинства организмов с половым размножением.Сборка синаптонемного комплекса начинается во время премейотической S-фазы и ранней мейотической профазы с образования единственной белковой оси, осевого элемента, вдоль двух сестринских хроматид каждой хромосомы. В ранней профазе аксиальный элемент, связанный с парой сестринских хроматид, тесно ассоциирует на ≈ ·100 нм по своей длине с аксиальным элементом гомологичной хромосомы; тогда они называются боковыми элементами.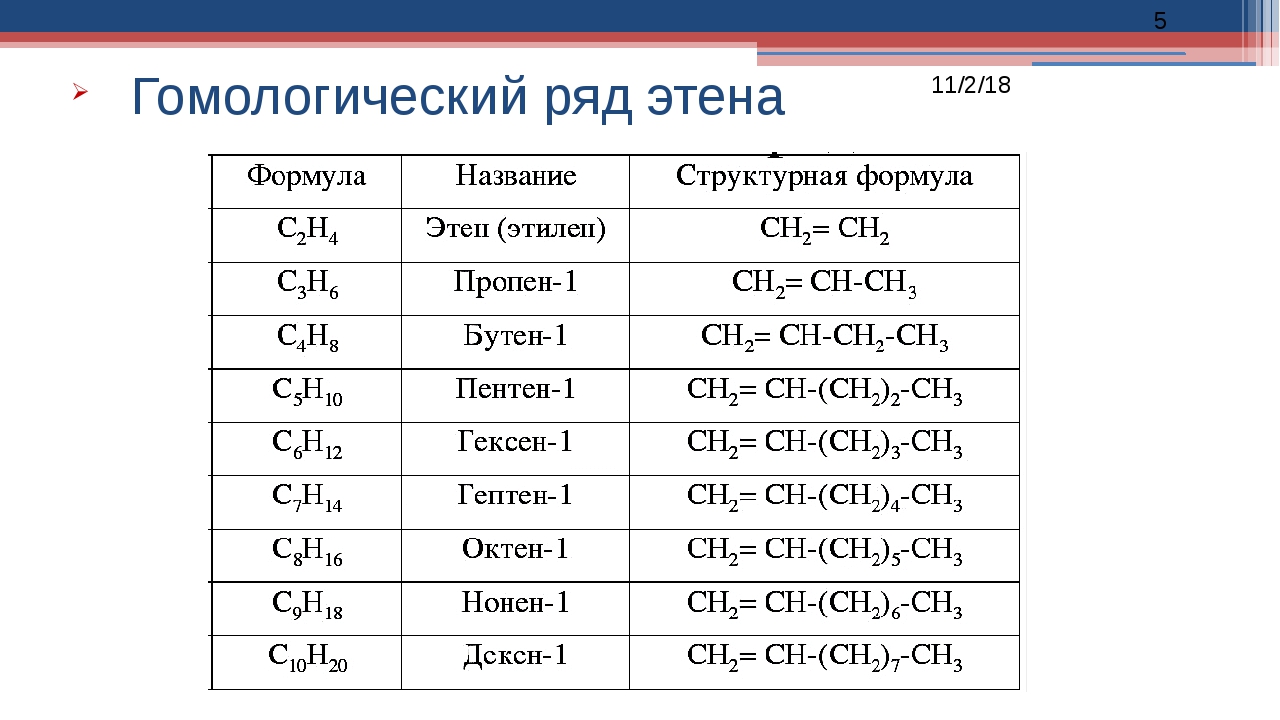 Продольное сближение гомологов опосредовано установкой многочисленных поперечных филаментов и образованием центрального элемента.Синаптонемный комплекс представляет собой лестничную структуру, образованную двумя боковыми элементами, поперечными филаментами и центральным элементом (рис. 2).
Продольное сближение гомологов опосредовано установкой многочисленных поперечных филаментов и образованием центрального элемента.Синаптонемный комплекс представляет собой лестничную структуру, образованную двумя боковыми элементами, поперечными филаментами и центральным элементом (рис. 2).
Компоненты синаптонемного комплекса идентифицированы у разных видов (обзоры [8, 9]). Белковый комплекс cohesin необходим для установления сцепления между сестринскими хроматидами во время репликации ДНК в соматических клетках. Cohesins являются компонентом аксиального/латерального элемента, но некоторые члены митотического комплекса заменены специфическими для мейоза паралогами.Конденсиновые белки также образуют часть аксиальных/латеральных элементов и необходимы для компактизации аксиальной длины и индивидуализации хромосом, сходно с их эффектами на митотическую конденсацию хромосом. Другие белки, такие как SYCP2 и SYCP3 у мыши, Hop1 и Red1 у почкующихся дрожжей, HIM-3 у C. elegans, Asy1 у Arabidopsis thaliana и ORD у Drosophila , являются членами латерального элемента. Помимо их роли в конденсации хроматина, аксиальные/латеральные элементы участвуют в выравнивании гомологов.Когезионная/конденсирующая хромосомная ось и/или ассоциированные белки, такие как Hop1p и HIM-3, участвуют в сборке поперечных филаментов. Регуляция способа, при котором DSB восстанавливаются до продуктов кроссинговера или некроссовера, также обеспечивается некоторыми белками латеральных элементов. Белки, образующие поперечные филаменты, идентифицированы у нескольких видов. К ним относятся Zip1p в S . cerevisiae , SYCP1 у млекопитающих, C(3)G у Drosophila , SYP-1 и SYP-2 у C . elegans, и ZYP1 в Arabidopsis . Эти белки имеют концевые глобулярные домены C и N, разделенные протяженной спиральной областью. Параллельные белковые димеры связываются через встречно-пальцевые N-концы и образуют тетрамер, который перекрывает промежуток между латеральными элементами.
elegans, Asy1 у Arabidopsis thaliana и ORD у Drosophila , являются членами латерального элемента. Помимо их роли в конденсации хроматина, аксиальные/латеральные элементы участвуют в выравнивании гомологов.Когезионная/конденсирующая хромосомная ось и/или ассоциированные белки, такие как Hop1p и HIM-3, участвуют в сборке поперечных филаментов. Регуляция способа, при котором DSB восстанавливаются до продуктов кроссинговера или некроссовера, также обеспечивается некоторыми белками латеральных элементов. Белки, образующие поперечные филаменты, идентифицированы у нескольких видов. К ним относятся Zip1p в S . cerevisiae , SYCP1 у млекопитающих, C(3)G у Drosophila , SYP-1 и SYP-2 у C . elegans, и ZYP1 в Arabidopsis . Эти белки имеют концевые глобулярные домены C и N, разделенные протяженной спиральной областью. Параллельные белковые димеры связываются через встречно-пальцевые N-концы и образуют тетрамер, который перекрывает промежуток между латеральными элементами. У мышей стабильные поперечные филаменты требуют ассоциации тетрамеров SYCP1 и SYCE1. Белки SYCE1 и SYCE2 располагаются в центральном элементе. Поперечные филаменты и белок центрального элемента необходимы для завершения перекрестного пути рекомбинационного взаимодействия.
У мышей стабильные поперечные филаменты требуют ассоциации тетрамеров SYCP1 и SYCE1. Белки SYCE1 и SYCE2 располагаются в центральном элементе. Поперечные филаменты и белок центрального элемента необходимы для завершения перекрестного пути рекомбинационного взаимодействия.
Взаимодействие между гомологичными хромосомами, упомянутое ранее, требует их расположения в непосредственной физической близости от ядра. Термин «выравнивание» обычно относится к топологическому расположению хромосом с параллельными плечами. Спаривание относится к тесным и стабильным взаимодействиям хромосом, которые включают несколько точек и приводят к тесной ассоциации гомологов по всей их длине. Хотя ранние исследования показали, что гомологичные хромосомы не спарены в лептотене [10], было предложено упорядоченное премейотическое интерфазное ядро с пресинаптическими гомологичными выравниваниями хромосом как часть этого порядка [11, 12].Однако исследования, проведенные на разных организмах, противоречат этому предположению. У грибов с зиготным мейозом, таких как Sordaria, Neurospora, и Coprinus , гомологичные хромосомы находятся в разных ядрах до оплодотворения. После кариогамии гомологи остаются разделенными до начала взаимодействия [13]. Среди растений окраску хромосом использовали для идентификации положения двух гомологов кукурузы, добавленных к овсем [14], а также двух гомологов ржи, добавленных к пшенице [15].Такие окрашенные пары хромосом занимают отдельные территории в большинстве ядер в премейотической интерфазе, что исключает роль премиотического расположения в выборе мейотического партнера. В последнее десятилетие все большее внимание привлекает пространственная организация генома млекопитающих в соматических клетках. Геном млекопитающих организован в виде структурированного ядра, в котором сворачивание и взаимное расположение хромосом составляет регуляторный механизм высокого порядка экспрессии генов [16]. Хромосомы млекопитающих организованы в дискретные, неперекрывающиеся хромосомные территории, причем хромосомы гомологов обычно не соседствуют [17].
У грибов с зиготным мейозом, таких как Sordaria, Neurospora, и Coprinus , гомологичные хромосомы находятся в разных ядрах до оплодотворения. После кариогамии гомологи остаются разделенными до начала взаимодействия [13]. Среди растений окраску хромосом использовали для идентификации положения двух гомологов кукурузы, добавленных к овсем [14], а также двух гомологов ржи, добавленных к пшенице [15].Такие окрашенные пары хромосом занимают отдельные территории в большинстве ядер в премейотической интерфазе, что исключает роль премиотического расположения в выборе мейотического партнера. В последнее десятилетие все большее внимание привлекает пространственная организация генома млекопитающих в соматических клетках. Геном млекопитающих организован в виде структурированного ядра, в котором сворачивание и взаимное расположение хромосом составляет регуляторный механизм высокого порядка экспрессии генов [16]. Хромосомы млекопитающих организованы в дискретные, неперекрывающиеся хромосомные территории, причем хромосомы гомологов обычно не соседствуют [17]. Расположение пары хромосом 1 человека и пары хромосом 8 мыши в сперматогониях указывает на отсутствие премейотического спаривания [18]. Только у таких организмов, как двукрылые, гомологичные хромосомы спариваются на протяжении всего жизненного цикла (соматическое спаривание). Поведение меченых хромосом с белком-репрессором GFP-lac во время мейоза самцов D. melanogaster указывает на то, что события спаривания, наблюдаемые в первом мейотическом делении, были продолжением премейотических спариваний, а не результатом механизмов, специфичных для мейоза [19].Таким образом, за исключением двукрылых, гомологичному спариванию способствуют механизмы, которые постепенно уменьшают физическое разделение между гомологами и различают гомологичные и негомологические взаимодействия, пока не завершатся тесным и стабильным соседством гомологов.
Расположение пары хромосом 1 человека и пары хромосом 8 мыши в сперматогониях указывает на отсутствие премейотического спаривания [18]. Только у таких организмов, как двукрылые, гомологичные хромосомы спариваются на протяжении всего жизненного цикла (соматическое спаривание). Поведение меченых хромосом с белком-репрессором GFP-lac во время мейоза самцов D. melanogaster указывает на то, что события спаривания, наблюдаемые в первом мейотическом делении, были продолжением премейотических спариваний, а не результатом механизмов, специфичных для мейоза [19].Таким образом, за исключением двукрылых, гомологичному спариванию способствуют механизмы, которые постепенно уменьшают физическое разделение между гомологами и различают гомологичные и негомологические взаимодействия, пока не завершатся тесным и стабильным соседством гомологов.
Способ, посредством которого гомологичные хромосомы сближаются в пространстве, узнают друг друга и вступают в стабильные взаимодействия, является одним из наиболее плохо изученных механизмов мейотического процесса. Раскрытие генетической и молекулярной основы поиска и обнаружения гомологии представляет собой цель многих текущих исследований различных организмов.В этой статье я сосредоточусь на клеточной активности, которая сближает гомологи и помогает обнаружить отличительные метки гомологии, кульминацией которых является образование гомологичных бивалентов.
Раскрытие генетической и молекулярной основы поиска и обнаружения гомологии представляет собой цель многих текущих исследований различных организмов.В этой статье я сосредоточусь на клеточной активности, которая сближает гомологи и помогает обнаружить отличительные метки гомологии, кульминацией которых является образование гомологичных бивалентов.
2. Управляемые теломерами движения хромосом, приближающиеся к гомологичным парам
Особенностью ядерной архитектуры в премейотической интерфазе, связанной с мейотическим спариванием, является распределение и ориентация центромер и теломер. В митотически активных клетках многих видов хромосомы сохраняют во время интерфазы географию предыдущей анафазы, т. е. конфигурацию Rabl, с центромерами, сосредоточенными на одной стороне ядра, и теломерами, развернутыми веером на противоположной стороне.Эта поляризация хромосом распространяется на премейотические клетки, особенно у видов с большими геномами [20], и гарантирует, что гомологичные сегменты, хотя и расположены на разделенных территориях, сохраняют одинаковые расстояния от ядерных полюсов, что сводит к минимуму перемещения хромосом при гомологичном спаривании [21]. .
.
У многих организмов теломеры всех хромосом инициируют неслучайное движение в начале мейоза, что приводит к их прогрессивному группированию, достигающему кульминации в плотный кластер.Эта супрахромосомная конфигурация, так называемый букет, консолидируется одновременно с инициацией выравнивания, спаривания и синапсов, особенно в областях, близких к концам хромосом, поэтому в настоящее время она рассматривается как мейотическая специфическая структура, участвующая в гомологичных взаимодействиях хромосом. 22–27]. Другие роли, включая разрешение двухвалентных блокировок и регуляцию рекомбинации, также были предложены для структуры букета [28]. Формирование букета является результатом трех взаимозависимых событий: прикрепления теломер к ядерной оболочке, кластеризации концов хромосом и движения теломер, опосредованного цитоскелетом (rev. [29]).
Молекулярные взаимодействия, ответственные за прикрепление концов хромосом к ядерной оболочке, требуют присутствия функциональных повторов теломер. Укорочение теломер у мышей с дефицитом теломеразы приводит к нарушению синапсов и снижению частоты рекомбинации [30]. Присутствие de novo теломерных повторов на центромерном конце телоцентрических хромосом, генерируемых неправильным делением центромер двуплечих хромосом, изменяет динамику центромеры. Эти повторы теломер оказывают цис-доминантный эффект на центромеру, которая может включаться в кластер теломер, в то время как исходные центромеры остаются на противоположном полюсе ядра [15].Когда теломерные последовательности расположены интеркалярно, как это происходит в кольцевой хромосоме кукурузы, они также связываются с другими теломерами и заставляют хромосому входить в букет [31].
Укорочение теломер у мышей с дефицитом теломеразы приводит к нарушению синапсов и снижению частоты рекомбинации [30]. Присутствие de novo теломерных повторов на центромерном конце телоцентрических хромосом, генерируемых неправильным делением центромер двуплечих хромосом, изменяет динамику центромеры. Эти повторы теломер оказывают цис-доминантный эффект на центромеру, которая может включаться в кластер теломер, в то время как исходные центромеры остаются на противоположном полюсе ядра [15].Когда теломерные последовательности расположены интеркалярно, как это происходит в кольцевой хромосоме кукурузы, они также связываются с другими теломерами и заставляют хромосому входить в букет [31].
Прикрепление теломер к внутренней поверхности ядерной оболочки требует наличия конститутивных белков, ассоциированных с концами хромосом, таких как Taz1, Rap1 и Rik1, и мейотических специфических белков, образующих мостик между теломерами и ядерной оболочкой. Эти белки включают Ndj1, Mps3 и Csm4 у почкующихся дрожжей [32, 33] и Sad1, Kms1, Bqt1 и Bqt2 у делящихся дрожжей [24, 34]. Некоторые из этих мейотических белков, такие как Sad1 и Kms1, являются трансмембранными белками с доменом SUN (Sad1-Unc-84) и доменом KASH (Klarsicht/ANC-1/Syne homology) соответственно [35]. Белок Sad1 ориентирован в нуклеоплазму и связан с теломерами комплексом Bqt1-Bqt2. Домен SUN белка Sad1, расположенный на его С-конце, как полагают, физически взаимодействует через межмембранное пространство с доменом KASH белка Kms1, интегрированным во внешнюю мембрану. Домен KASH соответствует короткой области С-конца белка, а большой N-концевой хвост простирается в цитоплазму и соединяет внешнюю мембрану с динеином, цитоплазматическим мотором микротрубочек, направленным на минус-конец.Мутации в Sad1 или Kms1 сильно уменьшают кроссинговер и нарушают сегрегацию гомологов, в то время как мутации в dynein вызывают более тонкие дефекты. Белки доменов SUN и KASH играют консервативную роль, устанавливая временные связи между концами хромосом и компонентами цитоскелета в интактной ядерной оболочке. Белок Mps3 почкующихся дрожжей является другим членом семейства белков домена SUN, который связан с теломерами с помощью Ndj1.
Некоторые из этих мейотических белков, такие как Sad1 и Kms1, являются трансмембранными белками с доменом SUN (Sad1-Unc-84) и доменом KASH (Klarsicht/ANC-1/Syne homology) соответственно [35]. Белок Sad1 ориентирован в нуклеоплазму и связан с теломерами комплексом Bqt1-Bqt2. Домен SUN белка Sad1, расположенный на его С-конце, как полагают, физически взаимодействует через межмембранное пространство с доменом KASH белка Kms1, интегрированным во внешнюю мембрану. Домен KASH соответствует короткой области С-конца белка, а большой N-концевой хвост простирается в цитоплазму и соединяет внешнюю мембрану с динеином, цитоплазматическим мотором микротрубочек, направленным на минус-конец.Мутации в Sad1 или Kms1 сильно уменьшают кроссинговер и нарушают сегрегацию гомологов, в то время как мутации в dynein вызывают более тонкие дефекты. Белки доменов SUN и KASH играют консервативную роль, устанавливая временные связи между концами хромосом и компонентами цитоскелета в интактной ядерной оболочке. Белок Mps3 почкующихся дрожжей является другим членом семейства белков домена SUN, который связан с теломерами с помощью Ndj1. Однако не было идентифицировано ни одного белка домена KASH, способного связать Mps3 с каким-либо компонентом цитоскелета.У C. elegans белок внутренней мембраны ядра SUN-1 взаимодействует с белком домена KASH ZYG-12 внешнего слоя, чтобы связать хромосомы через их центры спаривания с сетью микротрубочек и цитоплазматическим динеином [36]. У мутантов генов sun-1 или zyg-12 гомологичные хромосомы не спариваются должным образом и синапс беспорядочно. Белок внутренней ядерной мембраны SUN1 специфически связывается с теломерами между стадиями лептотены и диплотены во время профазы I мейоза у мышей [37].Разрушение sun1 предотвращает прикрепление теломер к ядерной оболочке, эффективное спаривание гомологов и синапсы. Однако о аналоге KASH для SUN1 до сих пор не сообщалось. У Arabidopsis двойных мутанта по генам sun1 и sun2 обнаруживают недостаточность синапсов и снижение частоты хиазм (JL Santos, личное сообщение).
Однако не было идентифицировано ни одного белка домена KASH, способного связать Mps3 с каким-либо компонентом цитоскелета.У C. elegans белок внутренней мембраны ядра SUN-1 взаимодействует с белком домена KASH ZYG-12 внешнего слоя, чтобы связать хромосомы через их центры спаривания с сетью микротрубочек и цитоплазматическим динеином [36]. У мутантов генов sun-1 или zyg-12 гомологичные хромосомы не спариваются должным образом и синапс беспорядочно. Белок внутренней ядерной мембраны SUN1 специфически связывается с теломерами между стадиями лептотены и диплотены во время профазы I мейоза у мышей [37].Разрушение sun1 предотвращает прикрепление теломер к ядерной оболочке, эффективное спаривание гомологов и синапсы. Однако о аналоге KASH для SUN1 до сих пор не сообщалось. У Arabidopsis двойных мутанта по генам sun1 и sun2 обнаруживают недостаточность синапсов и снижение частоты хиазм (JL Santos, личное сообщение).
Теломеры, прикрепленные к ядерной оболочке, сначала объединяются в ряд небольших агрегатов, которые, подвергаясь активным движениям, объединяются в плотный кластер.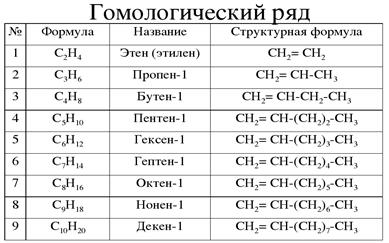 Ассоциация теломер и миграция теломер представляют собой два разных этапа консолидации букета, как следует из результатов, полученных при обработке премейотических и мейотических клеток пшеницы колхицином. Этот деполимеризующий микротрубочки агент ингибирует миграцию теломер и нарушает синапсы, но не влияет на ассоциацию теломер [38]. Эта ассоциация, скорее всего, возникает между теломерами, прикрепленными к очень близким участкам ядерной оболочки, как в случае концов гомологичных плеч изохромосомы.Эти плечи лежат близко друг к другу после последнего премейотического митоза и обнаруживают нормальный уровень спаривания и образования хиазм даже в присутствии колхицина [39].
Ассоциация теломер и миграция теломер представляют собой два разных этапа консолидации букета, как следует из результатов, полученных при обработке премейотических и мейотических клеток пшеницы колхицином. Этот деполимеризующий микротрубочки агент ингибирует миграцию теломер и нарушает синапсы, но не влияет на ассоциацию теломер [38]. Эта ассоциация, скорее всего, возникает между теломерами, прикрепленными к очень близким участкам ядерной оболочки, как в случае концов гомологичных плеч изохромосомы.Эти плечи лежат близко друг к другу после последнего премейотического митоза и обнаруживают нормальный уровень спаривания и образования хиазм даже в присутствии колхицина [39].
На входе в мейоз гомологичные хромосомы занимают пространственно разнесенные территории. Расстояние между гомологами может достигать нескольких мк м у организмов с большим геномом; например, ядро пшеницы в лептотене имеет диаметр ≈ 24 мкм м. Сокращение такого расстояния требует активного движения хромосом, которые, кроме того, должны быть правильно ориентированы, чтобы сблизить гомологичные хромосомы. Пространство между гомологами попадает в гораздо более уменьшенную шкалу у делящихся дрожжей с небольшим геномом (13,8 Mb) и тремя парами хромосом, включенными в клетку, имеющую размеры 3,5 µ м в диаметре и 10 µ м в диаметре. длина. В этом организме мейоз происходит в зиготе после слияния двух гаметных ядер, и гомологи должны быть заменены, чтобы выровняться. Выравнивание достигается характерными движениями удлиненного ядра, называемого ядром «конского хвоста». Ядро хвоща движется вперед и назад по зиготе в течение примерно 2 часов.Этот период, соответствующий профазе мейоза, предоставляет хромосомам единственную возможность спаривания и рекомбинации со своими гомологичными партнерами. Движения ядер опосредованы астральными микротрубочками, отходящими от тела полюса веретена (центр организации микротрубочек у грибов), и моторным белком динеином [40, 41]. Во время этого процесса белковый комплекс Sad1-Kms1 взаимодействует с теломерами на нуклеоплазматической стороне и с моторным белком динеином на цитоплазматической стороне.
Пространство между гомологами попадает в гораздо более уменьшенную шкалу у делящихся дрожжей с небольшим геномом (13,8 Mb) и тремя парами хромосом, включенными в клетку, имеющую размеры 3,5 µ м в диаметре и 10 µ м в диаметре. длина. В этом организме мейоз происходит в зиготе после слияния двух гаметных ядер, и гомологи должны быть заменены, чтобы выровняться. Выравнивание достигается характерными движениями удлиненного ядра, называемого ядром «конского хвоста». Ядро хвоща движется вперед и назад по зиготе в течение примерно 2 часов.Этот период, соответствующий профазе мейоза, предоставляет хромосомам единственную возможность спаривания и рекомбинации со своими гомологичными партнерами. Движения ядер опосредованы астральными микротрубочками, отходящими от тела полюса веретена (центр организации микротрубочек у грибов), и моторным белком динеином [40, 41]. Во время этого процесса белковый комплекс Sad1-Kms1 взаимодействует с теломерами на нуклеоплазматической стороне и с моторным белком динеином на цитоплазматической стороне. Таким образом, теломеры перемещаются под действием движущей силы, генерируемой динеиновым мотором на микротрубочках.Теломеры образуют группу рядом с телом полюса веретена, и это расположение вместе с колебательным движением ядер приводит к тому, что гомологи выровнены.
Таким образом, теломеры перемещаются под действием движущей силы, генерируемой динеиновым мотором на микротрубочках.Теломеры образуют группу рядом с телом полюса веретена, и это расположение вместе с колебательным движением ядер приводит к тому, что гомологи выровнены.
У почкующихся дрожжей сортировка и спаривание хромосом, происходящие в раннем мейозе, сопровождаются быстрыми теломерными движениями хромосом, при которых теломеры группируются возле тела полюса веретена, образуя букет. Эти движения обеспечиваются белками, прикрепленными к теломерам Ndj1 и Mps3, и белком Csm4, который участвует не в прикреплении ядерной оболочки к теломерам, а в передаче силы, генерируемой цитоскелетом [33].Направляемые теломерами движения хромосом у почкующихся дрожжей не зависят от комплексов динеин-микротрубочки [42]; они регулируются актиновыми филаментами, поскольку ингибирование полимеризации актина латрукулином В нарушает кластеризацию теломер [43]. На формирование букета влияет колхицин в мейоцитах растений. Однако движение теломер не опосредовано цитоплазматическими микротрубочками. В качестве мишени для колхицина был предложен ассоциированный с мембраной тубулин или родственные ему белки [44].
Однако движение теломер не опосредовано цитоплазматическими микротрубочками. В качестве мишени для колхицина был предложен ассоциированный с мембраной тубулин или родственные ему белки [44].
Принято считать, что формирование букета облегчает гомологичное спаривание за счет сокращения ядерного пространства, в котором хромосомы должны двигаться, чтобы найти гомологичного партнера [27].Формирование букета подразумевает, что все теломеры, независимо от их положения на периферии ядра, мигрируют в небольшой участок ядерной оболочки. Это движение, скорее всего, поляризовано положением тела полюса веретена у делящихся дрожжей. Однако в таких организмах, как растения, без определенного центра организации микротрубочек, трудно представить, как движение теломер ориентируется на конвергенцию в области, противоположной полюсу центромеры. Можно утверждать, что расположение Рабла способствует поляризованному движению.Однако силы конвергентного цитоскелета действуют по всей ядерной поверхности, поскольку они способны кластеризовать теломеры телоцентриков, расположенных на полюсе центромер, и двуплечих хромосом, расположенных в противоположном полушарии [15]. На подвижность теломер могут влиять нецитоплазматические факторы, такие как хромосомная конформация. Этот эффект стал очевидным, когда проследили миграцию отдельных концов двуплечей хромосомы в двух различных конформациях: субметацентрической хромосоме, то есть плечах хромосомы, имеющей очень разную длину, и метацентрической хромосоме, то есть плечах одинаковой длины.Ситуация с неравными плечами соответствует стандартной хромосоме 5R ржи, а метацентрическая конформация возникла в результате большой делеции, образовавшейся в длинном плече этой хромосомы. Короткое плечо одинаково в обеих конформациях хромосом, но его теломеры чаще достигают теломерного полюса при метацентрическом строении, чем при субметацентрическом строении [45]. Это изменчивое поведение указывает на то, что наличие длинного плеча оказывает некоторое сопротивление движению теломеры короткого плеча или препятствует ее прикреплению к ядерной мембране.
На подвижность теломер могут влиять нецитоплазматические факторы, такие как хромосомная конформация. Этот эффект стал очевидным, когда проследили миграцию отдельных концов двуплечей хромосомы в двух различных конформациях: субметацентрической хромосоме, то есть плечах хромосомы, имеющей очень разную длину, и метацентрической хромосоме, то есть плечах одинаковой длины.Ситуация с неравными плечами соответствует стандартной хромосоме 5R ржи, а метацентрическая конформация возникла в результате большой делеции, образовавшейся в длинном плече этой хромосомы. Короткое плечо одинаково в обеих конформациях хромосом, но его теломеры чаще достигают теломерного полюса при метацентрическом строении, чем при субметацентрическом строении [45]. Это изменчивое поведение указывает на то, что наличие длинного плеча оказывает некоторое сопротивление движению теломеры короткого плеча или препятствует ее прикреплению к ядерной мембране.
3. Перемещения хромосом, независимые от миграции теломер
Уровень гомологичного спаривания и синапсов, продуцируемых у мутантов с дефектом букета почкующихся дрожжей или кукурузы [25, 46], а также в мейоцитах, где образование букета ингибируется колхицином [38], указывает на что кластеризация теломер не является абсолютно необходимой для спаривания. Хотя кластеризация теломер может сокращать расстояние между гомологами и облегчать их распознавание, эта конвергентная сила влияет в основном на дистальные участки хромосом, особенно на большие хромосомы с длиной в зиготене порядка десятков или даже сотен мк м.Обширное спаривание крупных хромосом вида Allium предполагает, что силы притяжения между гомологами действуют в разных точках, равномерно распределенных по большей части длины хромосомы [47, 48]. Движения хромосом, которые делают возможными столкновения между интеркалярными областями, вероятно, являются результатом конформационных изменений хроматина, ведущих к удлинению хромосом (Рисунок 3), которые производятся одновременно с кластеризацией теломер при переходе лептотен-зиготена [15, 38].Хромосомы пшеницы и ржи умножают свою длину в 5 раз в лептотене по сравнению с премейотической интерфазой. Поскольку размер ядра остается прежним при переходе от лептотены к зиготене или даже уменьшается, разворачивание хроматина заставляет хромосомы перемещаться и охватывать все ядро.
Хотя кластеризация теломер может сокращать расстояние между гомологами и облегчать их распознавание, эта конвергентная сила влияет в основном на дистальные участки хромосом, особенно на большие хромосомы с длиной в зиготене порядка десятков или даже сотен мк м.Обширное спаривание крупных хромосом вида Allium предполагает, что силы притяжения между гомологами действуют в разных точках, равномерно распределенных по большей части длины хромосомы [47, 48]. Движения хромосом, которые делают возможными столкновения между интеркалярными областями, вероятно, являются результатом конформационных изменений хроматина, ведущих к удлинению хромосом (Рисунок 3), которые производятся одновременно с кластеризацией теломер при переходе лептотен-зиготена [15, 38].Хромосомы пшеницы и ржи умножают свою длину в 5 раз в лептотене по сравнению с премейотической интерфазой. Поскольку размер ядра остается прежним при переходе от лептотены к зиготене или даже уменьшается, разворачивание хроматина заставляет хромосомы перемещаться и охватывать все ядро. Это развертывание хроматина не ингибируется колхицином [38] и поэтому не зависит от миграции теломер. Движение, которое вызывает удлинение хромосом, может способствовать возникновению случайных столкновений между гомологичными хромосомами.Сокращение больших расстояний между гомологами во время зиготены особенно заметно у гетерозигот по инверсии, покрывающей почти 95% плеча хромосомы ржи. Наблюдения проводились на линии пшеницы с дисомным добавлением хромосомы 1R [49] и, следовательно, в ядрах диаметром более 20 мк мкм. Во время формирования букета субдистальная часть нормального плеча располагается на полюсе теломер, в то время как его гомологичный аналог в инвертированном плече расположен на полюсе центромеры.Однако во многих мейоцитах такие участки образуют пары и синапсы. Столкновения между отдаленными гомологичными областями могут возникать после удлинения хромосомы, как показано на рис. 4.
Это развертывание хроматина не ингибируется колхицином [38] и поэтому не зависит от миграции теломер. Движение, которое вызывает удлинение хромосом, может способствовать возникновению случайных столкновений между гомологичными хромосомами.Сокращение больших расстояний между гомологами во время зиготены особенно заметно у гетерозигот по инверсии, покрывающей почти 95% плеча хромосомы ржи. Наблюдения проводились на линии пшеницы с дисомным добавлением хромосомы 1R [49] и, следовательно, в ядрах диаметром более 20 мк мкм. Во время формирования букета субдистальная часть нормального плеча располагается на полюсе теломер, в то время как его гомологичный аналог в инвертированном плече расположен на полюсе центромеры.Однако во многих мейоцитах такие участки образуют пары и синапсы. Столкновения между отдаленными гомологичными областями могут возникать после удлинения хромосомы, как показано на рис. 4.
Визуализация ядерной динамики в мейоцитах кукурузы при жизни показала, что движения хромосом в зиготене включают повороты всего хроматина и движения отдельных сегментов хромосом [50]. ]. Не было установлено, были ли эти движения предназначены для содействия спариванию или были произведены после синапса.Однако предполагается, что движения отдельных сегментов хромосом происходят во время разворачивания хроматина. Кроме того, рекомбинация как таковой может увеличить подвижность хромосом. Быстрые движения хромосом, возникающие в ранней профазе у почкующихся дрожжей, модулируются рекомбинацией. Мутанты с дефектами рекомбинации не демонстрируют самых надежных и быстрых движений, характерных для дикого типа [33]. С другой стороны, в вегетативных клетках почкующихся дрожжей либо данный индуцированный ДР, либо несколько ДР, индуцированных γ -облучением, запускают подвижность всех хромосом как компонент механизма репарации [51, 52].DSB являются мощными триггерами реакции на повреждение ДНК, сложной сигнальной сети, участвующей в активации киназ контрольных точек клеточного цикла, которые останавливают цикл до тех пор, пока повреждение не будет восстановлено.
]. Не было установлено, были ли эти движения предназначены для содействия спариванию или были произведены после синапса.Однако предполагается, что движения отдельных сегментов хромосом происходят во время разворачивания хроматина. Кроме того, рекомбинация как таковой может увеличить подвижность хромосом. Быстрые движения хромосом, возникающие в ранней профазе у почкующихся дрожжей, модулируются рекомбинацией. Мутанты с дефектами рекомбинации не демонстрируют самых надежных и быстрых движений, характерных для дикого типа [33]. С другой стороны, в вегетативных клетках почкующихся дрожжей либо данный индуцированный ДР, либо несколько ДР, индуцированных γ -облучением, запускают подвижность всех хромосом как компонент механизма репарации [51, 52].DSB являются мощными триггерами реакции на повреждение ДНК, сложной сигнальной сети, участвующей в активации киназ контрольных точек клеточного цикла, которые останавливают цикл до тех пор, пока повреждение не будет восстановлено. Главным преобразователем сигнала DSB является ядерная протеинкиназа с мутацией атаксия-телеангиэктазия (ATM), которая фосфорилирует различные субстраты [53], что может приводить к изменениям в организации хроматина и динамике хромосом. Конформационные изменения хроматина, приводящие к удлинению хромосом в конце лептотены, не являются результатом ответа на повреждение ДНК, активируемого DSB, продуцируемыми Spo11; во время профазы I длина хромосом одинакова как у дикого типа, так и у нулевых мутантов spo11 у Arabidopsis и мыши [54, 55].
Главным преобразователем сигнала DSB является ядерная протеинкиназа с мутацией атаксия-телеангиэктазия (ATM), которая фосфорилирует различные субстраты [53], что может приводить к изменениям в организации хроматина и динамике хромосом. Конформационные изменения хроматина, приводящие к удлинению хромосом в конце лептотены, не являются результатом ответа на повреждение ДНК, активируемого DSB, продуцируемыми Spo11; во время профазы I длина хромосом одинакова как у дикого типа, так и у нулевых мутантов spo11 у Arabidopsis и мыши [54, 55].
4. Центромерные соединения
Функционально эквивалентные домены, отличные от теломер, присутствующие также во всех хромосомах, являются центромерами. Это области, участвующие в прикреплении хромосом к микротрубочкам и опосредующие движение хромосом во время клеточного деления. Обогащение различных повторяющихся последовательностей ДНК или белков, участвующих в их организации, может представлять собой возможный источник хромосомных взаимодействий при поиске гомологичного партнера. Эта возможная роль была подчеркнута с помощью пресинаптического спаривания центромер, наблюдаемого у широкого круга организмов, включая грибы, растения и животных (обзор [56]), особенно у аллополиплоидных пшениц [57, 58].Среди полиплоидных пшениц выделяют два уровня плоидности: тетраплоидный и гексаплоидный. Тетраплоидная пшеница, Triticum turgidum (макаронная пшеница), несет четыре набора из семи хромосом (формула генома AABB). Геномы A и B произошли от двух родственных диких диплоидных предшественников, T. urartu и Aegilops speltoides соответственно. Хлебная пшеница, T . aestivum — гексаплоидный вид (AABBDD), несущий геномы A и B тетраплоидной пшеницы и геном D диплоидного предка T.таущий . Несмотря на генетическое родство между хромосомами геномов А, В и D (гомеологичные хромосомы), мягкая пшеница образует 21 бивалент при диакинезе и метафазе I. Исключительное образование гомологичных бивалентов является результатом подавления гомеологического спаривания, главным образом контролируемого Ph2 (парный гомеологичный) локус на длинном плече хромосомы 5В [59, 60].
Эта возможная роль была подчеркнута с помощью пресинаптического спаривания центромер, наблюдаемого у широкого круга организмов, включая грибы, растения и животных (обзор [56]), особенно у аллополиплоидных пшениц [57, 58].Среди полиплоидных пшениц выделяют два уровня плоидности: тетраплоидный и гексаплоидный. Тетраплоидная пшеница, Triticum turgidum (макаронная пшеница), несет четыре набора из семи хромосом (формула генома AABB). Геномы A и B произошли от двух родственных диких диплоидных предшественников, T. urartu и Aegilops speltoides соответственно. Хлебная пшеница, T . aestivum — гексаплоидный вид (AABBDD), несущий геномы A и B тетраплоидной пшеницы и геном D диплоидного предка T.таущий . Несмотря на генетическое родство между хромосомами геномов А, В и D (гомеологичные хромосомы), мягкая пшеница образует 21 бивалент при диакинезе и метафазе I. Исключительное образование гомологичных бивалентов является результатом подавления гомеологического спаривания, главным образом контролируемого Ph2 (парный гомеологичный) локус на длинном плече хромосомы 5В [59, 60]. Сообщалось, что область размером 2,5 Mb, содержащая сегмент гетерохроматина, встроенный в кластер генов, родственных циклин-зависимой киназы Cdk2, представляет собой локус Ph2 [61].Однако механизм его действия еще предстоит выяснить.
Сообщалось, что область размером 2,5 Mb, содержащая сегмент гетерохроматина, встроенный в кластер генов, родственных циклин-зависимой киназы Cdk2, представляет собой локус Ph2 [61].Однако механизм его действия еще предстоит выяснить.
Зародышевые клетки пшеницы вступают в мейоз с центромерами, связанными в среднем попарно. Такое расположение предполагает, что центромеры участвуют в механизме сортировки хромосом и что образование гомологичных бивалентов начинается с премейотической ассоциации их центромер, стимулируемой Ph2 [57]. Идентификация центромер двух гомологичных хромосом ржи, добавленных к пшенице, показала, что центромерные ассоциации в основном негомологичны и не зависят от Ph2 [15].Центромеры группируются в более сложные структуры во время лептотены тетраплоидных и гексаплоидных пшениц. В некоторых мейоцитах обнаружено семь пресинаптических мультицентромерных комплексов. Это предложило гипотезу о том, что как гомологичные, так и гомеологичные центромеры, спаренные или непарные в премейотической интерфазе, становятся ассоциированными до синапса и что после распознавания гомологии под контролем Ph2 каждая мультицентромерная структура разрешается в пары гомологичных центромер [58]. . Эта гипотеза предполагает сильное увеличение гомологичной ассоциации центромер перед синапсисом и присутствие достаточных последовательностей ДНК, специфичных для хромосом, чтобы различать гомологов и гомеологов.Это кажется маловероятным, потому что центромеры обычно содержат обильные повторы, которые не уникальны для отдельных хромосом.
. Эта гипотеза предполагает сильное увеличение гомологичной ассоциации центромер перед синапсисом и присутствие достаточных последовательностей ДНК, специфичных для хромосом, чтобы различать гомологов и гомеологов.Это кажется маловероятным, потому что центромеры обычно содержат обильные повторы, которые не уникальны для отдельных хромосом.
Гомологичные центромеры ржи, присутствующие в различных пшенично-ржаных дисомных добавках, в пшенично-ржаных робертсоновских транслокациях или интрогрессированные в хромосомы пшеницы, приурочены к пространственно разделенным кластерам центромер в более чем 80% мейоцитов в начале лептотены [56]. Это происходит независимо от того, присутствовал ли Ph2 или нет. Частота ассоциации гомологичных центромер увеличивается с прогрессированием профазы I, что является результатом расширения синапсов.Таким образом, кластеризация пресинаптических центромер у пшеницы не основана на гомологии и, следовательно, не может способствовать распознаванию гомологичных хромосом. Это согласуется с тем, что на спаривание и рекомбинацию гомологичных плеч не влияет гетерозиготность по центромере, а также с тем фактом, что гомологичные центромеры не вызывают спаривания гомеологичных хромосом [62]. FISH-анализ поведения центромер и дистальных хромомеров в телоцентрических и двуплечих хромосомах подтвердил, что в поиске и распознавании гомологичного партнера участвуют не центромерные, а субтеломерные районы [62].
Это согласуется с тем, что на спаривание и рекомбинацию гомологичных плеч не влияет гетерозиготность по центромере, а также с тем фактом, что гомологичные центромеры не вызывают спаривания гомеологичных хромосом [62]. FISH-анализ поведения центромер и дистальных хромомеров в телоцентрических и двуплечих хромосомах подтвердил, что в поиске и распознавании гомологичного партнера участвуют не центромерные, а субтеломерные районы [62].
В мейозе почкующихся дрожжей дикого типа хромосомы объединяются в пары на своих центромерах независимо от хромосомной гомологии. Большинство центромерных пар являются негомологичными на стадии перед синапсисом, но подвергаются переключению до тех пор, пока все они не вовлекают гомологов [63]. Этот переход от случайного к гомологичному спариванию требует активности Spo11, указывающей на то, что смена партнера зависит от того же механизма, основанного на рекомбинации, который регулирует гомологичное спаривание и синапсис. Раннее негомологичное спаривание центромер зависит от белка синаптонемного комплекса Zip1, который связывает пространство между ядрами гомологичных хромосом, но оно не зависит от Zip2 и Zip3, которые являются компонентом латерального элемента синаптонемного комплекса. Гомологичное выравнивание не затрагивается у мутанта zip1 , хотя синаптонемный комплекс не способен застегиваться [64]. Это указывает на то, что начальные центромерные взаимодействия необязательны для гомологичного спаривания хромосом. Вполне вероятно, что и у дрожжей расширение синапсов за счет хромосомных плеч сближает гомологичные центромеры, которые первоначально были вовлечены в негомологичные пары. Это предположение согласуется с инициацией синапсов в местах перекрестной рекомбинации [65].Однако гомологичный синапсис инициируется также на центромерах после деполимеризации Zip1, соединяющего негомологичные ассоциации [66]. Молекулярную основу связи между негомологичными центромерами у пшеницы еще предстоит установить. Однако образование множественных ассоциаций при парном паттерне дрожжей можно объяснить различиями в размере области центромер. Большой размер центромеры пшеницы может способствовать множественным взаимодействиям. Пресинаптическая центромерная ассоциация не является важным компонентом механизма спаривания хромосом.
Гомологичное выравнивание не затрагивается у мутанта zip1 , хотя синаптонемный комплекс не способен застегиваться [64]. Это указывает на то, что начальные центромерные взаимодействия необязательны для гомологичного спаривания хромосом. Вполне вероятно, что и у дрожжей расширение синапсов за счет хромосомных плеч сближает гомологичные центромеры, которые первоначально были вовлечены в негомологичные пары. Это предположение согласуется с инициацией синапсов в местах перекрестной рекомбинации [65].Однако гомологичный синапсис инициируется также на центромерах после деполимеризации Zip1, соединяющего негомологичные ассоциации [66]. Молекулярную основу связи между негомологичными центромерами у пшеницы еще предстоит установить. Однако образование множественных ассоциаций при парном паттерне дрожжей можно объяснить различиями в размере области центромер. Большой размер центромеры пшеницы может способствовать множественным взаимодействиям. Пресинаптическая центромерная ассоциация не является важным компонентом механизма спаривания хромосом. Соответственно, у Drosophila , где гомологи спарены в начале мейоза, центромеры кластеризуются в ооцитах на ранней зиготене. Эта кластеризация центромер, по-видимому, определяет первый шаг в инициации синапсов [67].
Соответственно, у Drosophila , где гомологи спарены в начале мейоза, центромеры кластеризуются в ооцитах на ранней зиготене. Эта кластеризация центромер, по-видимому, определяет первый шаг в инициации синапсов [67].
В организмах, где гомологичное спаривание центромер управляется синапсом, были предложены другие потенциальные роли пресинаптической кластеризации центромер. Скопление центромер происходит одновременно с миграцией теломер и перемещением хромосом, возникающим в результате удлинения хромосом, происходящего во время лептотены и ранней зиготены.Закрепление хромосом с помощью этого механизма может обеспечить стабильность полюса центромер, уменьшить степень беспорядка, вносимого динамикой хромосом, и правильно направить конвергенцию теломер [62]. У Drosophila , человека и почкующихся дрожжей события нерасхождения во втором мейотическом делении обогащены центромерно-проксимальными кроссоверами [68-70]. Эти проксимальные кроссинговеры, вероятно, нарушают сцепление перицентромерной области, вызывая преждевременное разделение сестринских хроматид в анафазе I и случайное расхождение в анапахазе II. Ранняя кластеризация центромер может усиливать действие факторов, таких как гетерохроматин, который подавляет проксимальные кроссинговеры [71]. Наконец, предполагается, что ранняя негомологическая ассоциация центромер связана с образованием специфичных для мейоза кинетохор, которые располагаются бок о бок и коориентируются к одному и тому же полюсу в анафазе I [72].
Ранняя кластеризация центромер может усиливать действие факторов, таких как гетерохроматин, который подавляет проксимальные кроссинговеры [71]. Наконец, предполагается, что ранняя негомологическая ассоциация центромер связана с образованием специфичных для мейоза кинетохор, которые располагаются бок о бок и коориентируются к одному и тому же полюсу в анафазе I [72].
5. Механизмы гомологичного распознавания
5.1. Рекомбинационные взаимодействия между гомологами
Механизм, с помощью которого гомологичные хромосомы узнают друг друга и пары, различается у разных организмов.У многих эукариот гомологичное распознавание зависит от межхромосомных взаимодействий ДНК/ДНК, возникающих в результате начальных стадий репарации Spo11-индуцированных DSB. После резекции 5′-концов ДНК в местах разрывов полученные 3 одноцепочечные ДНК вторгаются в интактный дуплекс ДНК гомологичной хромосомы, что катализируется рекомбиназами Rad51 и Dmc1, и генерируют промежуточные соединения, способные оценивать гомологию. У некоторых грибов для гомологичного выравнивания требуется инициация рекомбинационных взаимодействий.У почкующихся дрожжей оценка гомологичного спаривания с помощью тестов Cre/ LoxP и FISH в диком типе, у одиночных мутантов spo11 и ndj1 и у двойных мутантов spo11 ndj1 дала значения спаривания. несколько ниже у мутанта ndj1 и намного ниже как у одиночного мутанта spo11 , так и у двойного мутанта spo11 ndj1 , чем у дикого типа. Отсутствие Spo11, независимо от того, сформирован букет или нет, оказывает одинаковый эффект на спаривание [73].Этот результат указывает на то, что мейотическая реорганизация теломер не является независимым от DSB компонентом механизма спаривания и оказывает влияние на гомологичное сопоставление посредством своей роли в мейотической рекомбинации. Электронно-микроскопический трехмерный анализ был использован в Sordaria , чтобы проследить пространственные отношения между хромосомами для всех стадий спаривания у дикого типа и мутантов [13].
У некоторых грибов для гомологичного выравнивания требуется инициация рекомбинационных взаимодействий.У почкующихся дрожжей оценка гомологичного спаривания с помощью тестов Cre/ LoxP и FISH в диком типе, у одиночных мутантов spo11 и ndj1 и у двойных мутантов spo11 ndj1 дала значения спаривания. несколько ниже у мутанта ndj1 и намного ниже как у одиночного мутанта spo11 , так и у двойного мутанта spo11 ndj1 , чем у дикого типа. Отсутствие Spo11, независимо от того, сформирован букет или нет, оказывает одинаковый эффект на спаривание [73].Этот результат указывает на то, что мейотическая реорганизация теломер не является независимым от DSB компонентом механизма спаривания и оказывает влияние на гомологичное сопоставление посредством своей роли в мейотической рекомбинации. Электронно-микроскопический трехмерный анализ был использован в Sordaria , чтобы проследить пространственные отношения между хромосомами для всех стадий спаривания у дикого типа и мутантов [13]. Гомологичные хромосомы выстраиваются в конце лептотены на расстоянии 400 нм до формирования букета.В отсутствие Spo11 гомологичные аксиальные элементы проявляют лишь редкие признаки узнавания и не образуют синаптонемных комплексов. Однако после того, как DSB были индуцированы облучением у нулевого мутанта spo11 , гомологи выравниваются. Количество парных сегментов связано с количеством DSB, измеренных фокусами Rad51, которые появляются в местах спаривания. Эти результаты показывают, что зависимое от рекомбинации гомологичное распознавание является важным компонентом механизма гомологичного спаривания.
Гомологичные хромосомы выстраиваются в конце лептотены на расстоянии 400 нм до формирования букета.В отсутствие Spo11 гомологичные аксиальные элементы проявляют лишь редкие признаки узнавания и не образуют синаптонемных комплексов. Однако после того, как DSB были индуцированы облучением у нулевого мутанта spo11 , гомологи выравниваются. Количество парных сегментов связано с количеством DSB, измеренных фокусами Rad51, которые появляются в местах спаривания. Эти результаты показывают, что зависимое от рекомбинации гомологичное распознавание является важным компонентом механизма гомологичного спаривания.
Быстрые перемещения хромосом, возникающие в профазе I мейоза у почкующихся дрожжей, требуют прикрепления теломер к ядерной оболочке и часто превышают 1 мк м/с. Хотя эти движения первоначально не зависят от рекомбинации, они качественно различны у мутантов, демонстрирующих дефекты рекомбинации, что позволяет предположить, что кинетика хромосом отвечает на молекулярные изменения в аппарате рекомбинации [33]. Мутанты в генах, которые связывают теломеры с цитоскелетом через интактную ядерную оболочку, обнаруживают постепенное изменение интенсивности движения хромосом.Скорость спаривания прямо коррелирует с силой двигательной активности, указывая на то, что быстрые движения хромосом способствуют гомологичным столкновениям и спариванию [74]. Это согласуется с тем фактом, что короткие хромосомы часто остаются несинапсированными, когда синапсы длинных хромосом закончились; ожидается, что взаимодействия будут происходить чаще между более длинными хромосомами.
Мутанты в генах, которые связывают теломеры с цитоскелетом через интактную ядерную оболочку, обнаруживают постепенное изменение интенсивности движения хромосом.Скорость спаривания прямо коррелирует с силой двигательной активности, указывая на то, что быстрые движения хромосом способствуют гомологичным столкновениям и спариванию [74]. Это согласуется с тем фактом, что короткие хромосомы часто остаются несинапсированными, когда синапсы длинных хромосом закончились; ожидается, что взаимодействия будут происходить чаще между более длинными хромосомами.
Динамика, лежащая в основе гомологичного поиска, была исследована in vivo с использованием двух меченых гомологичных локусов в вегетативных клетках почкующихся дрожжей [51].Эти два локуса занимают в основном отдельные области ядра и перемещаются независимо друг от друга в замкнутом пространстве, составляющем 2,7% объема ядра при отсутствии повреждений ДНК. После индукции DSB эти два локуса спариваются в десять раз чаще. В разрезанной хромосоме наблюдается сильное увеличение подвижности локуса, который может исследовать ядерный объем в десять раз больший. Подвижность сигнала в неразрезанной хромосоме также увеличивается; этот локус исследует ядерный объем в четыре раза больше. Увеличение подвижности хромосом зависит от рекомбиназы Rad51 и от Sae2, белка, очень рано участвующего в процессинге концов ДНК.Эти результаты поддерживают модель гомологичного спаривания, основанную на активации механизма поиска гомологии путем обнаружения DSB. Во время поиска гомологии сломанная молекула перемещается в ядро, проверяя гомологию среди последовательностей, с которыми она сталкивается, пока не найдет партнера. Активация механизма поиска гомологии значительно увеличивает область ядра, которую необходимо исследовать посредством взаимодействий ДНК/ДНК. Сходный механизм может быть активирован Spo11-индуцированными DSB в мейотических клетках. Несколько DSB, репарированных вдоль каждой гомологичной пары, приведут к их продольному сближению.Считается, что молекулярные события рекомбинации происходят в цитологических структурах, называемых узелками рекомбинации.
Подвижность сигнала в неразрезанной хромосоме также увеличивается; этот локус исследует ядерный объем в четыре раза больше. Увеличение подвижности хромосом зависит от рекомбиназы Rad51 и от Sae2, белка, очень рано участвующего в процессинге концов ДНК.Эти результаты поддерживают модель гомологичного спаривания, основанную на активации механизма поиска гомологии путем обнаружения DSB. Во время поиска гомологии сломанная молекула перемещается в ядро, проверяя гомологию среди последовательностей, с которыми она сталкивается, пока не найдет партнера. Активация механизма поиска гомологии значительно увеличивает область ядра, которую необходимо исследовать посредством взаимодействий ДНК/ДНК. Сходный механизм может быть активирован Spo11-индуцированными DSB в мейотических клетках. Несколько DSB, репарированных вдоль каждой гомологичной пары, приведут к их продольному сближению.Считается, что молекулярные события рекомбинации происходят в цитологических структурах, называемых узелками рекомбинации. Количество и распределение узелков ранней рекомбинации согласуются с этим способом распознавания гомологии [75].
Количество и распределение узелков ранней рекомбинации согласуются с этим способом распознавания гомологии [75].
У почкующихся дрожжей движения, генерируемые или усиливаемые мейотическими DSB, скорее всего, имеют достаточную величину, чтобы вызывать столкновения, обмены нитями и спаривание между гомологичными сайтами, первоначально расположенными в любом положении в ядре. Ожидается, что у эукариот с большими геномами и ядерными объемами в десять и более раз таких движений будет недостаточно для завершения спаривания между всеми гомологичными парами, поэтому гомологи выравниваются и инициируют синапсис после формирования букета [56].Эволюционно законсервированная особенность мейотического букета состоит в том, что его организация не зависит от рекомбинации, поскольку теломеры кластеризуются у мутантов мышей, почкующихся дрожжей или Sordaria , лишенных Spo11 [76–78]. У почкующихся дрожжей и Sordaria прикрепление теломер к ядерной оболочке необходимо для создания динамики хромосом, способной обеспечить гомологичную идентификацию до консолидации букета. У многих высших эукариот формирование букета уменьшает большое расстояние между гомологами, что делает возможной их идентификацию.Это было продемонстрировано в случае пары хромосом ржи 5R, присутствующей в аддитивной линии пшеницы-5R. В то время как генетический фон пшеницы дает нормальную частоту хиазм, поведение хромосомы 5R обусловлено ее субметацентрической конформацией. Миграция теломер плеча 5RS при формировании букета часто бывает задержанной или неполной [45]. За этой аномальной динамикой теломер (Рисунок 5) следует отказ в спаривании, образовании синапсов и хиазм в соответствующей паре плеч, что демонстрирует, что кластеризация теломер повышает эффективность механизма гомологичного поиска для обеспечения мейотического спаривания.
У многих высших эукариот формирование букета уменьшает большое расстояние между гомологами, что делает возможной их идентификацию.Это было продемонстрировано в случае пары хромосом ржи 5R, присутствующей в аддитивной линии пшеницы-5R. В то время как генетический фон пшеницы дает нормальную частоту хиазм, поведение хромосомы 5R обусловлено ее субметацентрической конформацией. Миграция теломер плеча 5RS при формировании букета часто бывает задержанной или неполной [45]. За этой аномальной динамикой теломер (Рисунок 5) следует отказ в спаривании, образовании синапсов и хиазм в соответствующей паре плеч, что демонстрирует, что кластеризация теломер повышает эффективность механизма гомологичного поиска для обеспечения мейотического спаривания.
У многих видов растений синапсис начинается в местах, близких к концам, за которым следуют дополнительные интеркалярные инициации для завершения сборки синаптонемного комплекса. Хиазмы также распределены неслучайно и располагаются в дистальной половине хромосом. У пшеницы исследования с использованием делеционных линий позволили предположить, что хиазмы возникают в основном в регионах с высокой плотностью генов [79, 80]. То, что не положение, а последовательность ДНК или организация хроматина, обычно присутствующая в дистальной части данного плеча хромосомы, определяет образование кроссинговера, было продемонстрировано на пшенице и ржи с использованием инверсий, которые изменяют положение богатых кроссинговером областей от дистальный к проксимальному.Паттерн хиазм инвертирован у инверсионных гомозигот [81, 82]. У инверсионных гетерозигот динамика нормального и инвертированного плеч во время зиготены демонстрирует, что богатые кроссинговером области более активны в распознавании гомологичного партнера и развитии синапсов, чем регионы с плохим кроссинговером [49]. У нормальных гомозигот области, богатые кроссинговером, расположены вблизи концов хромосом, и их ассоциация облегчается кластеризацией теломер. У инверсионных гетерозигот области, богатые кроссинговером, располагаются в центре одной хромосомы и дистально в гомологе, и то же самое происходит с областями, бедными кроссинговером.
У пшеницы исследования с использованием делеционных линий позволили предположить, что хиазмы возникают в основном в регионах с высокой плотностью генов [79, 80]. То, что не положение, а последовательность ДНК или организация хроматина, обычно присутствующая в дистальной части данного плеча хромосомы, определяет образование кроссинговера, было продемонстрировано на пшенице и ржи с использованием инверсий, которые изменяют положение богатых кроссинговером областей от дистальный к проксимальному.Паттерн хиазм инвертирован у инверсионных гомозигот [81, 82]. У инверсионных гетерозигот динамика нормального и инвертированного плеч во время зиготены демонстрирует, что богатые кроссинговером области более активны в распознавании гомологичного партнера и развитии синапсов, чем регионы с плохим кроссинговером [49]. У нормальных гомозигот области, богатые кроссинговером, расположены вблизи концов хромосом, и их ассоциация облегчается кластеризацией теломер. У инверсионных гетерозигот области, богатые кроссинговером, располагаются в центре одной хромосомы и дистально в гомологе, и то же самое происходит с областями, бедными кроссинговером. Однако регионы, богатые кроссовером, чаще соединяются друг с другом. Вполне вероятно, что динамика, вызванная удлинением хромосом, облегчает столкновения между гомологичными сайтами двух антипараллельных плеч хромосом, но результаты спаривания указывают на более высокую подвижность областей, богатых кроссинговером. Возникает вопрос, связана ли внутрихромосомная дифференциация в способности находить совпадения с паттерном ДР. Однако данные о распределении ДЦР в хромосомах пшеницы и ржи отсутствуют.Карта мейотических DSB с высоким разрешением по всему геному была построена только у двух эукариот, S. cerevisiae [83] и мышей [84]. У обоих видов карта DSB показывает разумное согласие с картой перекрестного распределения.
Однако регионы, богатые кроссовером, чаще соединяются друг с другом. Вполне вероятно, что динамика, вызванная удлинением хромосом, облегчает столкновения между гомологичными сайтами двух антипараллельных плеч хромосом, но результаты спаривания указывают на более высокую подвижность областей, богатых кроссинговером. Возникает вопрос, связана ли внутрихромосомная дифференциация в способности находить совпадения с паттерном ДР. Однако данные о распределении ДЦР в хромосомах пшеницы и ржи отсутствуют.Карта мейотических DSB с высоким разрешением по всему геному была построена только у двух эукариот, S. cerevisiae [83] и мышей [84]. У обоих видов карта DSB показывает разумное согласие с картой перекрестного распределения.
5.2. Нерекомбинационные взаимодействия между гомологами
Drosophila и C. elegans должны иметь независимые от рекомбинации механизмы, которые приводят к сортировке и спариванию хромосом, поскольку нормальный гомологичный синапсис возникает у мутантов, блокирующих образование DSB [85, 86]. Хромосомные перестройки C. elegans показали, что каждая хромосома содержит цис-действующую последовательность, расположенную вблизи одного конца, которая способствует гомологичной идентификации и необходима для гомологичной рекомбинации и сегрегации [87]. Эти сайты называются центрами спаривания и взаимодействуют с семейством из четырех паралогичных белков, каждый из которых содержит два атипичных цинковых пальца C2h3. Каждый белок локализуется в центрах спаривания одной или двух пар хромосом во время ранней мейотической профазы: ZIM-1 на хромосомах II и III, ZIM-2 на хромосоме V, ZIM-3 на хромосомах I и IV и HIM-8 на X хромосомах. хромосома.Мотивы короткой последовательности, обогащенные соответствующим цис-действующим элементом, селективно привлекают эти белки in vivo . Вставки кластера рекрутинговых мотивов в хромосому, лишенную эндогенного центра спаривания, достаточно для восстановления гомологичного спаривания, синапсов, перекрестной рекомбинации и сегрегации. Было предложено проверять гомологию на уникальных последовательностях, перемежающихся и/или примыкающих к основным кластерам сайтов связывания на каждой хромосоме.
Хромосомные перестройки C. elegans показали, что каждая хромосома содержит цис-действующую последовательность, расположенную вблизи одного конца, которая способствует гомологичной идентификации и необходима для гомологичной рекомбинации и сегрегации [87]. Эти сайты называются центрами спаривания и взаимодействуют с семейством из четырех паралогичных белков, каждый из которых содержит два атипичных цинковых пальца C2h3. Каждый белок локализуется в центрах спаривания одной или двух пар хромосом во время ранней мейотической профазы: ZIM-1 на хромосомах II и III, ZIM-2 на хромосоме V, ZIM-3 на хромосомах I и IV и HIM-8 на X хромосомах. хромосома.Мотивы короткой последовательности, обогащенные соответствующим цис-действующим элементом, селективно привлекают эти белки in vivo . Вставки кластера рекрутинговых мотивов в хромосому, лишенную эндогенного центра спаривания, достаточно для восстановления гомологичного спаривания, синапсов, перекрестной рекомбинации и сегрегации. Было предложено проверять гомологию на уникальных последовательностях, перемежающихся и/или примыкающих к основным кластерам сайтов связывания на каждой хромосоме. Хотя C. elegans не образует букет, движения хромосом, которые сводят вместе гомологичные центры спаривания, генерируются механизмом, сходным с механизмом кластеризации теломер.Эти движения опосредуются белковым комплексом SUN-1/ZYG-12 (SUN-KASH), который охватывает ядерную оболочку и соединяет каждый центр спаривания с моторным белком микротрубочек dynein [88]. Комплекс, образованный центром спаривания и трансмембранными белками SUN-1 и ZYG-12, перемещает хромосомы вдоль ядерной оболочки с помощью динеин-зависимых сил микротрубочек.
Хотя C. elegans не образует букет, движения хромосом, которые сводят вместе гомологичные центры спаривания, генерируются механизмом, сходным с механизмом кластеризации теломер.Эти движения опосредуются белковым комплексом SUN-1/ZYG-12 (SUN-KASH), который охватывает ядерную оболочку и соединяет каждый центр спаривания с моторным белком микротрубочек dynein [88]. Комплекс, образованный центром спаривания и трансмембранными белками SUN-1 и ZYG-12, перемещает хромосомы вдоль ядерной оболочки с помощью динеин-зависимых сил микротрубочек.
Другой сайт спаривания был идентифицирован в половых хромосомах Drosophila [89] . Этот центр спаривания содержит 200–250 тандемных повторов генов рРНК (также называемых рДНК) и расположен в гетерохроматине Х-хромосомы и вблизи основания короткого плеча Y-хромосомы.Делеция рДНК Х-хромосомы вызывает нарушение спаривания X-Y и нерасхождение половых хромосом. Эти мейотические аномалии частично восстанавливаются у трансгенных самцов при вставке одной копии рДНК в делетированную Х-хромосому. Активность спаривания заключается в последовательности из 240 п.н., которая повторяется в тандемных массивах от шести до десяти копий в межгенном спейсере, обнаруженном между рибосомными генами. Эти 240-пн повторы функционируют для рекрутирования двух белков, SNM и MNM, которые необходимы для стабильного спаривания гомологов и сегрегации у самцов.Взаимодействия между белками половой хромосомы, по-видимому, замещают хиазмы в ахиазматном мужском мейозе. Однако сайты связывания белков SNM и MNM на аутосомах остаются неопределенными. Аутосомы у обоих полов и два XX у самок уже спарены на входе в мейоз. Как достигается это расположение, было исследовано с помощью высокопроизводительной технологии FISH, которая сделала возможным полногеномный скрининг РНКи на факторы, участвующие в соматическом спаривании. Были идентифицированы как гены, способствующие спариванию, так и гены, препятствующие спариванию, что подтверждает идею о том, что гомологичное спаривание опосредуется балансом факторов с противоположными функциями [90].
Активность спаривания заключается в последовательности из 240 п.н., которая повторяется в тандемных массивах от шести до десяти копий в межгенном спейсере, обнаруженном между рибосомными генами. Эти 240-пн повторы функционируют для рекрутирования двух белков, SNM и MNM, которые необходимы для стабильного спаривания гомологов и сегрегации у самцов.Взаимодействия между белками половой хромосомы, по-видимому, замещают хиазмы в ахиазматном мужском мейозе. Однако сайты связывания белков SNM и MNM на аутосомах остаются неопределенными. Аутосомы у обоих полов и два XX у самок уже спарены на входе в мейоз. Как достигается это расположение, было исследовано с помощью высокопроизводительной технологии FISH, которая сделала возможным полногеномный скрининг РНКи на факторы, участвующие в соматическом спаривании. Были идентифицированы как гены, способствующие спариванию, так и гены, препятствующие спариванию, что подтверждает идею о том, что гомологичное спаривание опосредуется балансом факторов с противоположными функциями [90]. Группа промоторов спаривания включает множество генов с функциями, связанными с митотическим делением клеток, в то время как гены, участвующие в S-фазе прогрессии, репликации ДНК и компактизации хроматина, включены в группу антиспаривания. Молекулярная основа механизма гомологичного распознавания и то, как спаривание способствует или препятствует возникновению пар, представляет собой захватывающую задачу для будущих исследований.
Группа промоторов спаривания включает множество генов с функциями, связанными с митотическим делением клеток, в то время как гены, участвующие в S-фазе прогрессии, репликации ДНК и компактизации хроматина, включены в группу антиспаривания. Молекулярная основа механизма гомологичного распознавания и то, как спаривание способствует или препятствует возникновению пар, представляет собой захватывающую задачу для будущих исследований.
Другой механизм обнаружения гомологии, в котором РНК опосредует гомологичную ассоциацию хромосом во время мейоза, был описан в недавнем сообщении у делящихся дрожжей [91].Расположение флуоресцентно меченого локуса sme2 отслеживали в живых мейотических клетках. Ген sme2 кодирует некодирующие полиаденилированные РНК мейРНК-S и мейРНК-L, которые позволяют клеткам переключаться с митоза на мейоз за счет связывания белков Mei2 и Mmi1. Mei2 является положительным регулятором мейотического входа, который образует отчетливую точку в локусе sme2 на хромосоме II в профазном ядре хвоща. Mmi1 связывается с мейотически важными мРНК и дестабилизирует их в митотических клетках.В зиготических клетках Mmi1 рекрутируется в сайт sme2 и инактивируется Mei2, что позволяет прогрессировать мейозу. Две точки Mei2, первоначально присутствующие в зиготических клетках, соединяются друг с другом сразу после кариогамии и остаются парными в течение большей части стадии хвоща. Надежное спаривание в локусе sme2 происходит в отсутствие рекомбинации и требует транскрипции мейРНК-L длиной 1,5 т.п.н. Активность спаривания локализуется на 3′-конце мейРНК-L-транскрипта, и для ее эффективности требуются РНК-транскрипты с обеих хромосом.РНК-опосредованное спаривание в локусе sme2 указывает на то, что РНК-содержащие комплексы, распространяющиеся вдоль хромосом, могут действовать как специфичные для хромосом идентификаторы. Хотя нет доказательств, подтверждающих роль РНК в мейотическом спаривании у других организмов, представление о том, что транскрипты, заякоренные в одной хромосоме, могут быть использованы для идентификации гомологичных последовательностей, представляется возможным [92].
Mmi1 связывается с мейотически важными мРНК и дестабилизирует их в митотических клетках.В зиготических клетках Mmi1 рекрутируется в сайт sme2 и инактивируется Mei2, что позволяет прогрессировать мейозу. Две точки Mei2, первоначально присутствующие в зиготических клетках, соединяются друг с другом сразу после кариогамии и остаются парными в течение большей части стадии хвоща. Надежное спаривание в локусе sme2 происходит в отсутствие рекомбинации и требует транскрипции мейРНК-L длиной 1,5 т.п.н. Активность спаривания локализуется на 3′-конце мейРНК-L-транскрипта, и для ее эффективности требуются РНК-транскрипты с обеих хромосом.РНК-опосредованное спаривание в локусе sme2 указывает на то, что РНК-содержащие комплексы, распространяющиеся вдоль хромосом, могут действовать как специфичные для хромосом идентификаторы. Хотя нет доказательств, подтверждающих роль РНК в мейотическом спаривании у других организмов, представление о том, что транскрипты, заякоренные в одной хромосоме, могут быть использованы для идентификации гомологичных последовательностей, представляется возможным [92]. Контакты между гомологичными хромосомами, центрированные на DSB, продуцируемых активными генами, были продемонстрированы в соматических клетках человека в G0/G1-фазе [93].Эти контакты аннулируются ингибиторами транскрипции актиномицином D и α-аманитином, что указывает на то, что транскрипция кодирующей РНК может опосредовать гомологичное распознавание.
Контакты между гомологичными хромосомами, центрированные на DSB, продуцируемых активными генами, были продемонстрированы в соматических клетках человека в G0/G1-фазе [93].Эти контакты аннулируются ингибиторами транскрипции актиномицином D и α-аманитином, что указывает на то, что транскрипция кодирующей РНК может опосредовать гомологичное распознавание.
Другие хромосомные особенности, отличные от рекомбинационных взаимодействий ДНК/ДНК или РНК-опосредованного спаривания, были предложены для участия в гомологичном распознавании. Так обстоит дело с характером распределения когезина в осевых элементах непарных мейотических хромосом у мышей [94]. Две специфичные для мейоза субъединицы, Rec8 и Rad21L, cohesin располагаются на осевых элементах в ранних мейотических клетках.Комплексы, содержащие Rec8 или Rad21L, пространственно разделены, и характер их распределения вдоль оси хромосом одинаков у двух гомологов. Эти результаты привели к предположению, что когезиновая композиция осевых элементов может обеспечить код для гомологичного распознавания.
6. Коррекция спаривания
Исследовательские перемещения хромосом в окружающем пространстве в поисках партнера генерируют взаимодействия между гомологами, а также между негомологами, что поддерживается негомологичным синапсом, продуцируемым у ряда мейотических мутантов в разных организмах.Это подразумевает, что механизм идентификации гомологии должен включать некоторый компонент с функцией дестабилизации и диссоциации негомологических столкновений, чтобы повторно инициировать дополнительные попытки поиска, которые завершаются гомологичным спариванием. Мало что известно о способе, с помощью которого хромосомы обнаруживают и устраняют нежелательные взаимодействия. Недавнее сообщение показало своевременное одновременное исчезновение неправильных хромосомных ассоциаций с дезорганизацией кластера теломер [49]. У гетерозигот при почти полной инверсии плеча хромосомы ржи неправильные и нестабильные ассоциации, включающие терминальные или проксимальные области нормальных и инвертированных хромосом, исчезают одновременно с растворением кластеров теломер. Между тем дистальные и проксимальные гомологичные области продолжают взаимодействовать и подвергаться стабильному синапсу. Это свидетельствует о том, что в коррекции спаривания участвуют хромосомные движения, которые рассеивают концы хромосом при растворении букета. Такие движения могут раздвигать негомологичные хромосомы, которые инициируют новый поиск правильного партнера. Различие между стабильными и нестабильными ассоциациями и, следовательно, консолидацией или исправлением спаривания, по-видимому, зависит от способности хромосомных контактов образовывать кроссовер или нет.
Между тем дистальные и проксимальные гомологичные области продолжают взаимодействовать и подвергаться стабильному синапсу. Это свидетельствует о том, что в коррекции спаривания участвуют хромосомные движения, которые рассеивают концы хромосом при растворении букета. Такие движения могут раздвигать негомологичные хромосомы, которые инициируют новый поиск правильного партнера. Различие между стабильными и нестабильными ассоциациями и, следовательно, консолидацией или исправлением спаривания, по-видимому, зависит от способности хромосомных контактов образовывать кроссовер или нет.
Растворение кластеризации теломер, скорее всего, связано с коррекцией спаривания и синапсов, производимых у автотриплоидов и автотетраплоидов тутового шелкопряда Bombyx mori , а также у аллополиплоидных пшениц. Самки B. mori подвергаются хиазматному мейозу, в то время как самцы образуют хиазмы и демонстрируют стандартный мейотический путь. У триплоидных самок гомологичные хромосомы образуют триваленты в зиготене, которые к концу пахитены превращаются в биваленты плюс униваленты. Аналогичная синаптическая коррекция происходит у тетраплоидных самок; каждый квадривалент, образующийся в зиготене, превращается в два бивалента в пахитене. Однако у аутотриплоидных и аутотетраплоидных самцов мультивалентность сохраняется до метафазы I [95–97]. У аллополиплоидных пшениц хромосомные взаимодействия, возникающие в ранней зиготене, приводят к обнаружению гомологии между гомологами, а также между гомологами, что приводит к возникновению синаптического паттерна, характеризующегося наличием поливалентных конфигураций синаптонемных комплексов, образованных как гомологами, так и гомеологами, в дополнение к гомологичным бивалентам. 98–100].Такие мультиваленты восстанавливаются до гомологичных бивалентов в поздней зиготе и пахитене. Этот механизм коррекции спаривания находится под контролем гомеологического супрессора спаривания Ph2. В присутствии Ph2 хиазмы образуются только между гомологичными хромосомами, но при отсутствии Ph2 хиазмы могут образовываться и между гомеологами, а мультиваленты сохраняются до метафазы I [101, 102].
Аналогичная синаптическая коррекция происходит у тетраплоидных самок; каждый квадривалент, образующийся в зиготене, превращается в два бивалента в пахитене. Однако у аутотриплоидных и аутотетраплоидных самцов мультивалентность сохраняется до метафазы I [95–97]. У аллополиплоидных пшениц хромосомные взаимодействия, возникающие в ранней зиготене, приводят к обнаружению гомологии между гомологами, а также между гомологами, что приводит к возникновению синаптического паттерна, характеризующегося наличием поливалентных конфигураций синаптонемных комплексов, образованных как гомологами, так и гомеологами, в дополнение к гомологичным бивалентам. 98–100].Такие мультиваленты восстанавливаются до гомологичных бивалентов в поздней зиготе и пахитене. Этот механизм коррекции спаривания находится под контролем гомеологического супрессора спаривания Ph2. В присутствии Ph2 хиазмы образуются только между гомологичными хромосомами, но при отсутствии Ph2 хиазмы могут образовываться и между гомеологами, а мультиваленты сохраняются до метафазы I [101, 102]. Таким образом, хиазмы обусловливают также динамику спаривания и синапсов у полиплоидных пшениц и тутового шелкопряда.
Таким образом, хиазмы обусловливают также динамику спаривания и синапсов у полиплоидных пшениц и тутового шелкопряда.
Молекулярная основа дезорганизации букета плохо изучена. В мейозе почкующихся дрожжей отсутствие когезиновой субъединицы Rec8 ингибирует выход из кластеризации теломер и нарушает колокализацию тела полюса веретена и кластера теломер [103]. Экспрессия митотического cohesin компонента SCC1 из промотора rec8 в мейотических клетках восстанавливает эти дефекты, указывая на то, что cohesin опосредует ассоциацию кластера теломер и тела полюса веретена и выход из стадии букета.С др. стороны, теломер-обусловленные быстрые движения хромосом, которые, как считается, способствуют гомологичному спариванию [74], также были предложены в качестве строгого фактора для устранения нежелательных связей у почкующихся дрожжей [104]. Соответственно в C . elegans , движения хромосом, опосредованные комплексом SUN-1/ZYG-12, которые объединяют гомологичные сайты спаривания, также сотрудничают, чтобы ингибировать инициацию синапсов между временно ассоциированными негомологичными центрами.