Законы Менделя — Википедия
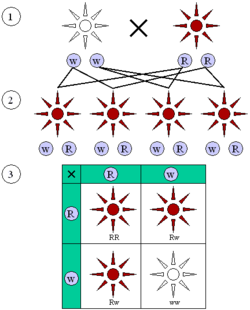 Схема первого и второго закона Менделя. 1) Растение с белыми цветками (две копии рецессивного аллеля w) скрещивается с растением с красными цветками (две копии доминантного аллеля R). 2) У всех растений-потомков цветы красные и одинаковый генотип Rw. 3) При самооплодотворении у 3/4 растений второго поколения цветки красные (генотипы RR + 2Rw) и у 1/4 — белые (ww).
Схема первого и второго закона Менделя. 1) Растение с белыми цветками (две копии рецессивного аллеля w) скрещивается с растением с красными цветками (две копии доминантного аллеля R). 2) У всех растений-потомков цветы красные и одинаковый генотип Rw. 3) При самооплодотворении у 3/4 растений второго поколения цветки красные (генотипы RR + 2Rw) и у 1/4 — белые (ww).Законы Менделя — принципы передачи наследственных признаков от родительских организмов к их потомкам, вытекающие из экспериментов Грегора Менделя. Эти принципы послужили основой для классической генетики и впоследствии были объяснены как следствие молекулярных механизмов наследственности. Хотя в русскоязычных учебниках обычно описывают три закона, «первый закон» открыт не Менделем. Особое значение из открытых Менделем закономерностей имеет «гипотеза чистоты гамет»
В начале XIX века Джон Госс (John Goss), экспериментируя с горохом, показал, что при скрещивании растений с зеленовато-голубыми горошинами и с желтовато-белыми в первом поколении получались жёлто-белые. Однако, при втором поколении, не проявляющиеся у гибридов первого поколения, и названные позже Менделем рецессивными признаки вновь проявлялись, причём растения с ними не давали расщепление при самоопылении[1].
Огюстен Сажрэ (фр., 1763—1851), французский растениевод, проводил эксперименты по гибридизации тыквенных, главным образом дынь. Сажрэ впервые в истории гибридизации стал изучать отдельные признаки скрещивающихся растений (мякоть, кожура и т. д.). Он установил, что при гибридизации родительские признаки распределяются между потомками без всякого смешения между собой. Таким образом, Сажрэ пришёл к установлению решающего свойства наследственности: в своей статье «Соображения об образовании гибридов, вариант и разновидностей» (1825 г.) он указывал на наличие наследственности «константной» вместо «слитной» наследственности [2].
Шарль Ноден (1815—1899), скрещивая различные виды дурмана, обнаружил преобладание признаков дурмана Datura tatula над Datura stramonium, причём это не зависело от того, какое растение материнское, а какое — отцовское[1].
Таким образом, к середине XIX века было открыто явление доминантности, единообразие гибридов в первом поколении (все гибриды первого поколения похожи друг на друга), расщепление и комбинаторику признаков во втором поколении. Тем не менее, Мендель, высоко оценивая работы предшественников, указывал, что всеобщего закона образования и развития гибридов ими не было найдено, и их опыты не обладают достаточной достоверностью для определения численных соотношений. Нахождение такого достоверного метода и математический анализ результатов, которые помогли создать теорию наследственности, является главной заслугой Менделя[1]
- Мендель изучал, как наследуются отдельные признаки.
- Мендель выбрал из всех признаков только альтернативные — такие, которые имели у его сортов два чётко различающихся варианта (семена либо гладкие, либо морщинистые; промежуточных вариантов не бывает). Такое сознательное сужение задачи исследования позволило чётко установить общие закономерности наследования.
- Мендель спланировал и провёл масштабный эксперимент. Им было получено от семеноводческих фирм 34 сорта гороха, из которых он отобрал 22 «чистых» (не дающих расщепления по изучаемым признакам при самоопылении) сорта. Затем он проводил искусственную гибридизацию сортов, а полученные гибриды скрещивал между собой. Он изучил наследование семи признаков, изучив в общей сложности около 20 000 гибридов второго поколения. Эксперимент облегчался удачным выбором объекта: горох в норме — самоопылитель, но на нём легко проводить искусственную гибридизацию.
- Мендель одним из первых в биологии использовал точные количественные методы для анализа данных. На основе знания теории вероятностей он понял необходимость анализа большого числа скрещиваний для устранения роли случайных отклонений.
Закон единообразия гибридов первого поколения[править | править код]
Проявление у гибридов признака только одного из родителей Мендель назвал доминированием.
Закон единообразия гибридов первого поколения (первый закон Менделя) — при скрещивании двух гомозиготных организмов, относящихся к разным чистым линиям и отличающихся друг от друга по одной паре альтернативных проявлений признака, всё первое поколение гибридов (F1) окажется единообразным и будет нести проявление признака одного из родителей[3].
Этот закон также известен как «закон доминирования признаков». Его формулировка основывается на понятии чистой линии относительно исследуемого признака — на современном языке это означает гомозиготность особей по этому признаку. Понятие гомозиготности было введено позднее У. Бэтсоном в 1902 году
При скрещивании чистых линий гороха с пурпурными цветками и гороха с белыми цветками Мендель заметил, что взошедшие потомки растений были все с пурпурными цветками, среди них не было ни одного белого. Мендель не раз повторял опыт, использовал другие признаки. Если он скрещивал горох с жёлтыми и зелёными семенами, у всех потомков семена были жёлтыми. Если он скрещивал горох с гладкими и морщинистыми семенами, у потомства были гладкие семена. Потомство от высоких и низких растений было высоким.
Итак, гибриды первого поколения всегда единообразны по данному признаку и приобретают признак одного из родителей. Этот признак — более сильный, доминантный (термин введён Менделем от латинского dominus), всегда подавлял другой, рецессивный[3].
Кодоминирование и неполное доминирование[править | править код]
Некоторые противоположные признаки находятся не в отношении полного доминирования (когда один всегда подавляет другой у гетерозиготных особей), а в отношении неполного доминирования. Например, при скрещивании чистых линий львиного зева с пурпурными и белыми цветками особи первого поколения имеют розовые цветки. При скрещивании чистых линий андалузских кур чёрной и белой окраски в первом поколении рождаются куры серой окраски. При неполном доминировании гетерозиготы имеют признаки, промежуточные между признаками рецессивной и доминантной гомозигот.
При кодоминировании, в отличие от неполного доминирования, у гетерозигот признаки проявляются одновременно (смешанно). Типичный пример кодоминирования — наследование групп крови системы АВ0 у человека, где А и В — доминантные гены, а 0 — рецессивный. По этой системе генотип 00 определяет первую группу крови, АА и А0 — вторую, ВВ и В0 — третью, а АВ будет определять четвёртую группу крови. Т.о. всё потомство людей с генотипами АА (вторая группа) и ВВ (третья группа) будет иметь генотип АВ (четвёртая группа). Их фенотип не является промежуточным между фенотипами родителей, так как на поверхности эритроцитов присутствуют оба агглютиногена (А и В).
Явления кодоминирования и неполного доминирования признаков слегка видоизменяет первый закон Менделя: «Гибриды первого поколения от скрещивания чистых линий особей с противоположными признаками всегда одинаковы по этому признаку: проявляют доминирующий признак, если признаки находятся в отношении доминирования, или смешанный (промежуточный) признак, если они находятся в отношении кодоминирования (неполного доминирования)».
Закон расщепления (второй закон Менделя) — при скрещивании двух гетерозиготных потомков первого поколения между собой, во втором поколении наблюдается расщепление в определённом числовом отношении: по фенотипу 3:1, по генотипу 1:2:1.
Скрещивание организмов двух чистых линий, различающихся по проявлениям одного изучаемого признака, за которые отвечают аллели одного гена, называется моногибридное скрещивание.
Явление, при котором скрещивание гетерозиготных особей приводит к образованию потомства, часть которого несёт доминантный признак, а часть — рецессивный, называется расщеплением. Следовательно, расщепление — это распределение доминантных и рецессивных признаков среди потомства в определённом числовом соотношении. Рецессивный признак у гибридов первого поколения не исчезает, а только подавляется и проявляется во втором гибридном поколении.
Объяснение[править | править код]
Закон чистоты гамет — в каждую гамету попадает только один аллель из пары аллелей данного гена родительской особи.
В норме гамета всегда чиста от второго гена аллельной пары. Этот факт, который во времена Менделя не мог быть твердо установлен, называют также гипотезой чистоты гамет. В дальнейшем эта гипотеза была подтверждена цитологическими наблюдениями. Из всех закономерностей наследования, установленных Менделем, данный «Закон» носит наиболее общий характер (выполняется при наиболее широком круге условий).
Гипотеза чистоты гамет. Мендель предположил, что при образовании гибридов наследственные факторы не смешиваются, а сохраняются в неизменном виде. У гибрида присутствуют оба фактора — доминантный и рецессивный, но проявление признака определяет доминантный наследственный фактор, рецессивный же подавляется. Связь между поколениями при половом размножении осуществляется через половые клетки — гаметы. Следовательно, необходимо допустить, что каждая гамета несет только один фактор из пары. Тогда при оплодотворении слияние двух гамет, каждая из которых несет рецессивный наследственный фактор, будет приводить к образованию организма с рецессивным признаком, проявляющимся фенотипически. Слияние же гамет, каждая из которых несет доминантный фактор, или же двух гамет, одна из которых содержит доминантный, а другая рецессивный фактор, будет приводить к развитию организма с доминантным признаком. Таким образом, появление во втором поколении рецессивного признака одного из родителей может быть только при двух условиях: 1) если у гибридов наследственные факторы сохраняются в неизменном виде; 2) если половые клетки содержат только один наследственный фактор из аллельной пары. Расщепление потомства при скрещивании гетерозиготных особей Мендель объяснил тем, что гаметы генетически чисты, то есть несут только один ген из аллельной пары. Гипотезу (теперь её называют законом) чистоты гамет можно сформулировать следующим образом: при образовании половых клеток в каждую гамету попадает только один аллель из пары аллелей данного гена.
Известно, что в каждой клетке организма в большинстве случаев имеется совершенно одинаковый диплоидный набор хромосом. Две гомологичные хромосомы обычно содержат каждая по одному аллелю данного гена. Генетически «чистые» гаметы образуются следующим образом:
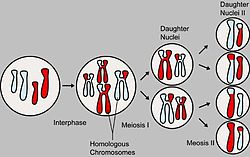 Основные этапы мейоза
Основные этапы мейозаНа схеме показан мейоз клетки с диплоидным набором 2n=4 (две пары гомологичных хромосом). Отцовские и материнские хромосомы обозначены разным цветом.
В процессе образования гамет у гибрида гомологичные хромосомы во время I мейотического деления попадают в разные клетки. При слиянии мужских и женских гамет получается зигота с диплоидным набором хромосом. При этом половину хромосом зигота получает от отцовского организма, половину — от материнского. По данной паре хромосом (и данной паре аллелей) образуются два сорта гамет. При оплодотворении гаметы, несущие одинаковые или разные аллели, случайно встречаются друг с другом. В силу статистической вероятности при достаточно большом количестве гамет в потомстве 25 % генотипов будут гомозиготными доминантными, 50 % — гетерозиготными, 25 % — гомозиготными рецессивными, то есть устанавливается отношение 1АА:2Аа:1аа (расщепление по генотипу 1:2:1). Соответственно по фенотипу потомство второго поколения при моногибридном скрещивании распределяется в отношении 3:1 (3/4 особей с доминантным признаком, 1/4 особей с рецессивным). Таким образом, при моногибридном скрещивании цитологическая основа расщепления признаков — расхождение гомологичных хромосом и образование гаплоидных половых клеток в мейозе.
Закон независимого наследования признаков[править | править код]
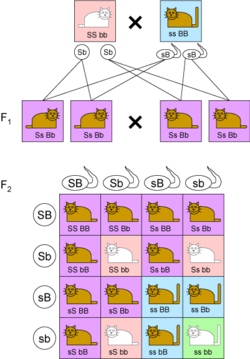 Иллюстрация независимого наследования признаков
Иллюстрация независимого наследования признаковОпределение[править | править код]
Закон независимого наследования (третий закон Менделя) — при скрещивании двух особей, отличающихся друг от друга по двум (и более) парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях (как и при моногибридном скрещивании).
Когда скрещивались гомозиготные растения, отличающиеся по нескольким признакам, таким как белые и пурпурные цветы и желтые или зелёные горошины, наследование каждого из признаков следовало первым двум законам, и в потомстве они комбинировались таким образом, как будто их наследование происходило независимо друг от друга. Первое поколение после скрещивания обладало доминантным фенотипом по всем признакам. Во втором поколении наблюдалось расщепление фенотипов по формуле 9:3:3:1, то есть 9:16 были с пурпурными цветами и желтыми горошинами, 3:16 с белыми цветами и желтыми горошинами, 3:16 с пурпурными цветами и зелёными горошинами, 1:16 с белыми цветами и зелёными горошинами.
Объяснение[править | править код]
Менделю попались признаки, гены которых находились в разных парах гомологичных хромосом (нуклеопротеидных структур в ядре эукариотической клетки, в которых сосредоточена бо́льшая часть наследственной информации и которые предназначены для её хранения, реализации и передачи) гороха. При мейозе гомологичные хромосомы разных пар комбинируются в гаметах случайным образом. Если в гамету попала отцовская хромосома первой пары, то с равной вероятностью в эту гамету может попасть как отцовская, так и материнская хромосома второй пары. Поэтому признаки, гены которых находятся в разных парах гомологичных хромосом, комбинируются независимо друг от друга. (Впоследствии выяснилось, что из исследованных Менделем семи пар признаков у гороха, у которого диплоидное число хромосом 2n=14, гены, отвечающие за одну из пар признаков, находились в одной и той же хромосоме. Однако Мендель не обнаружил нарушения закона независимого наследования, так как сцепления между этими генами не наблюдалось из-за большого расстояния между ними).
Основные положения теории наследственности Менделя[править | править код]
В современной интерпретации эти положения следующие:
- За наследственные признаки отвечают дискретные (отдельные, не смешивающиеся) наследственные факторы — гены (термин «ген» предложен в 1909 г. В.Иогансеном).
- Каждый диплоидный организм содержит пару аллелей данного гена, отвечающих за данный признак; один из них получен от отца, другой — от матери.
- Наследственные факторы передаются потомкам через половые клетки. При формировании гамет в каждую из них попадает только по одному аллелю из каждой пары (гаметы «чисты» в том смысле, что не содержат второго аллеля).
В соответствии с законами Менделя наследуются только моногенные признаки. Если за фенотипический признак отвечает более одного гена (а таких признаков абсолютное большинство), он имеет более сложный характер наследования.
Условия выполнения закона расщепления при моногибридном скрещивании[править | править код]
Расщепление 3 : 1 по фенотипу и 1 : 2 : 1 по генотипу выполняется приближенно и лишь при следующих условиях:
- Изучается большое число скрещиваний (большое число потомков).
- Гаметы, содержащие аллели А и а, образуются в равном числе (обладают равной жизнеспособностью).
- Нет избирательного оплодотворения: гаметы, содержащие любой аллель, сливаются друг с другом с равной вероятностью.
- Зиготы (зародыши) с разными генотипами одинаково жизнеспособны.
- Родительские организмы принадлежат к чистым линиям, то есть действительно гомозиготны по изучаемому гену (АА и аа).
- Признак действительно моногенный
- Признак не сцеплен с половыми хромосомами
Условия выполнения закона независимого наследования[править | править код]
- Все условия, необходимые для выполнения закона расщепления.
- Расположение генов, отвечающих за изучаемые признаки, в разных парах хромосом (несцепленность).
Условия выполнения закона чистоты гамет[править | править код]
- Нормальный ход мейоза. В результате нерасхождения хромосом в одну гамету могут попасть обе гомологичные хромосомы из пары. В этом случае гамета будет нести по паре аллелей всех генов, которые содержатся в данной паре хромосом.
- Гайсинович А.Е. Зарождение и развитие генетики. — М.: Наука, 1988. — 424 с. — ISBN 5-02-005265-5.
- Дубинин Н. П. Общая генетика. — М.: «Наука», 1986. — 560 с.
- В.И. Иванов, Н.В. Барышникова, Дж. С. Билева. Генетика / Под ред. В.И. Иванова. — М.: Академкнига, 2007. — 638 с. — 2000 экз. — ISBN 978-5-94628-288-8.
Законы Менделя, подготовка к ЕГЭ по биологии
В предыдущей статье мы познакомились с фундаментальными понятиями и методами генетики. Настало время их применить при изучении нового раздела — Менделевской генетики, основанной на законах, открытых Грегором Менделем.
Мендель следовал некоторым принципам в своих исследованиях, которые привели его работы к успеху:
- Использовал гибридологический метод генетики, подвергая скрещиванию растения гороха с четко различающимися признаками: желтый — зеленый цвет семян, гладкая — морщинистая форма семян
- Учитывал при скрещивании не всю совокупность признаков, а отдельные альтернативные признаки (желтый — зеленый цвет семян)
- Вел количественный учет потомков в ряду поколений, анализировал потомство каждой особи
- При размножении использовал чистые линии — группы растений, которые генетически однородны (гомозиготы AA, aa) и потомки которых не имеют разнообразия по изучаемому признаку

Введем несколько новых терминов, которые нам пригодятся. Скрещивание может быть:
- Моногибридным — в случае если скрещиваемые особи отличаются только по одному исследуемому признаку (цвет семян)
- Дигибридным — если скрещиваемые особи отличаются по двум различным признакам (цвет и форма семян)
В схеме решения генетическое задачи есть некоторые обозначения: ♀ — женский организм, ♂ — мужской организм, P — родительские организмы, F1 — гибриды первого поколения, F2 — гибриды второго поколения. Вероятно, имеет смысл сохранить картинку ниже себе на гаджет, если вы только приступаете к изучению генетики 😉
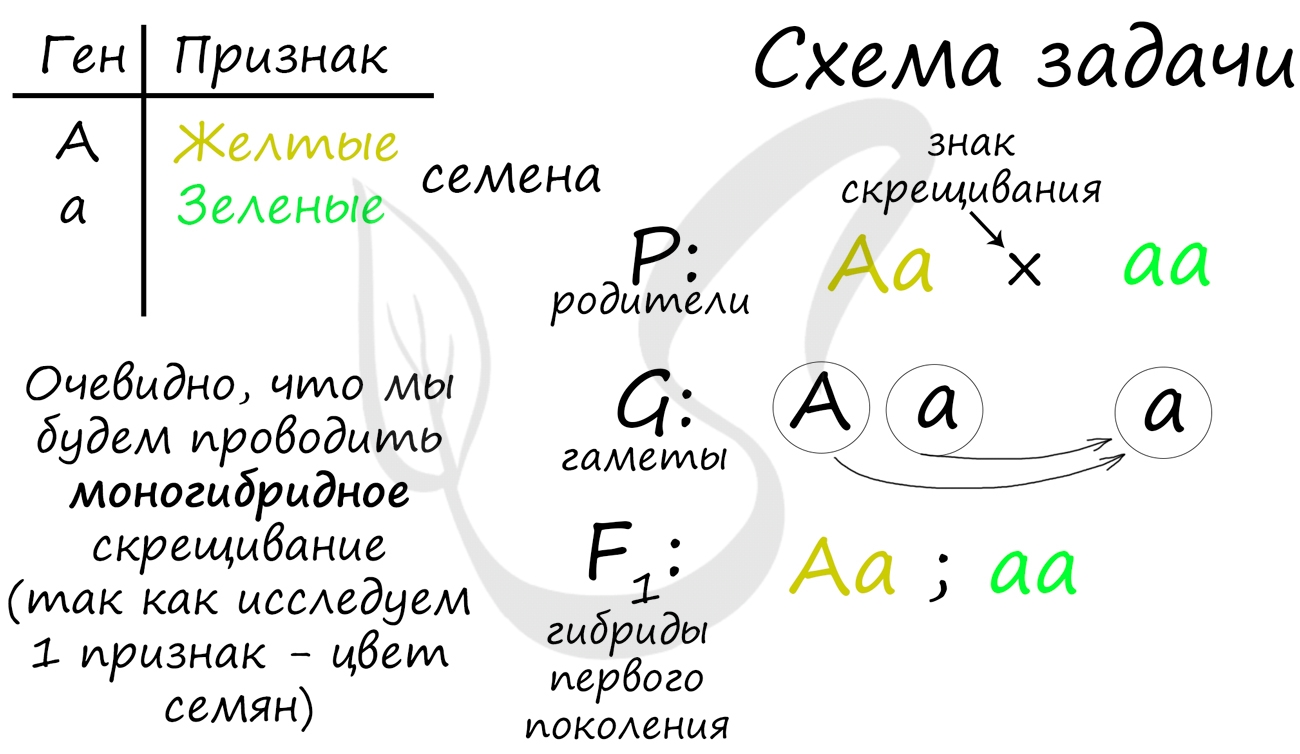
Спешу сообщить вам, что браки между людьми (в отличие от насильственного скрещивания гороха) происходят только по любви и взаимному согласию! Поэтому в задачах, где речь идет о людях, не следует ставить знак скрещивания «×» между родительскими особями. В таком случае ставьте знак «→» — «стрелу Амура», чтобы привести в восхищение экзаменатора 🙂
Первый закон Менделя — закон единообразия
С него часто начинаются генетические задачи (в качестве первого скрещивания). Этот закон гласит о том, что при скрещивании гомозиготных особей, отличающихся одной или несколькими парами альтернативных признаков, все гибриды первого поколения будут единообразны по данным признакам.
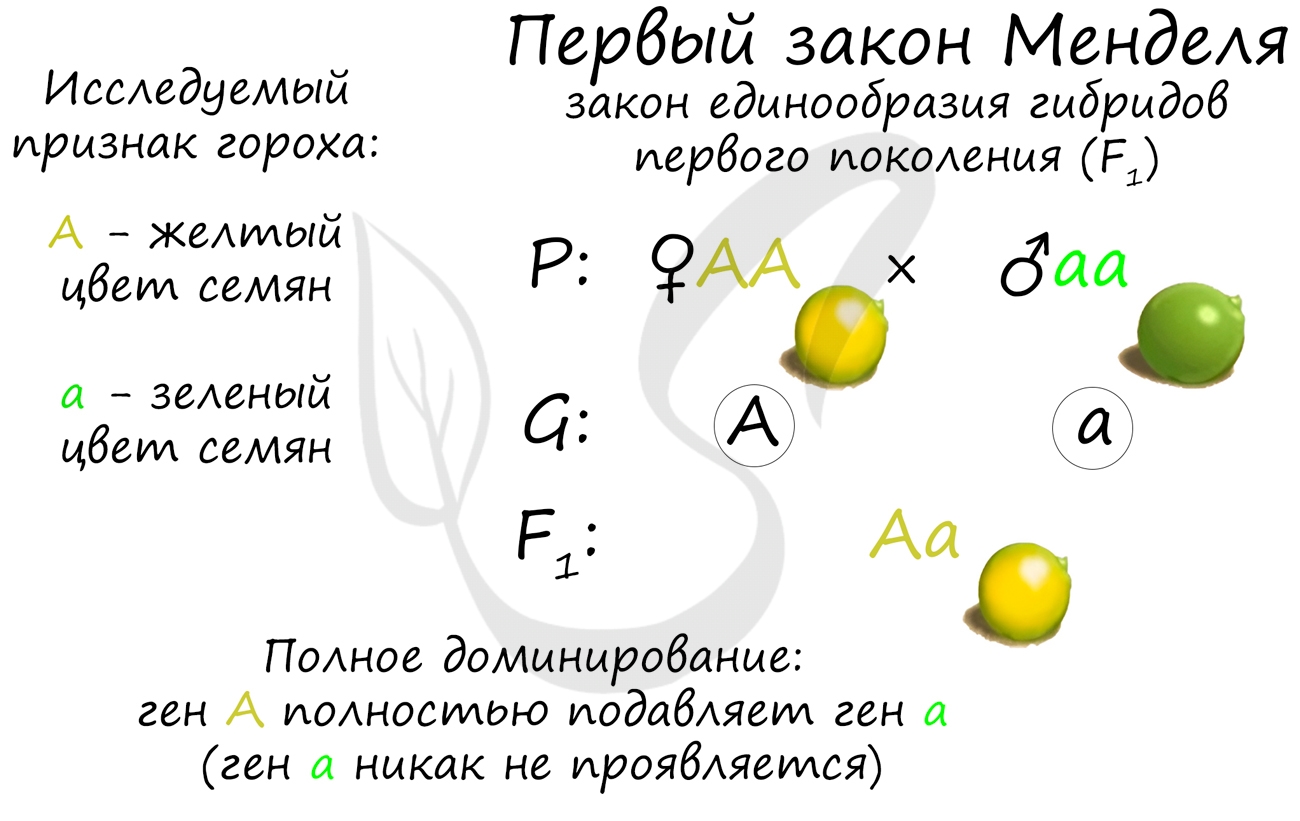
Этот закон основан на варианте взаимодействия между генами — полном доминировании. При таком варианте один ген — доминантный, полностью подавляет другой ген — рецессивный. В эксперименте, который мы только что изучили, Мендель скрещивал чистые линии гороха с желтыми (АА) и зелеными (aa) семенами, в результате все потомство имело желтый цвет семян (Aa) — было единообразно.
Анализирующее скрещивание
Часто генотип особи не изучен и представляет загадку. Как быть генетику в данном случае? Иногда проще всего применить анализирующее скрещивание — скрещивание гибридной особи (у которой не известен генотип) с гомозиготой по рецессивному признаку.
Анализируя полученное потомство, можно сделать вывод о генотипе гибридной особи.
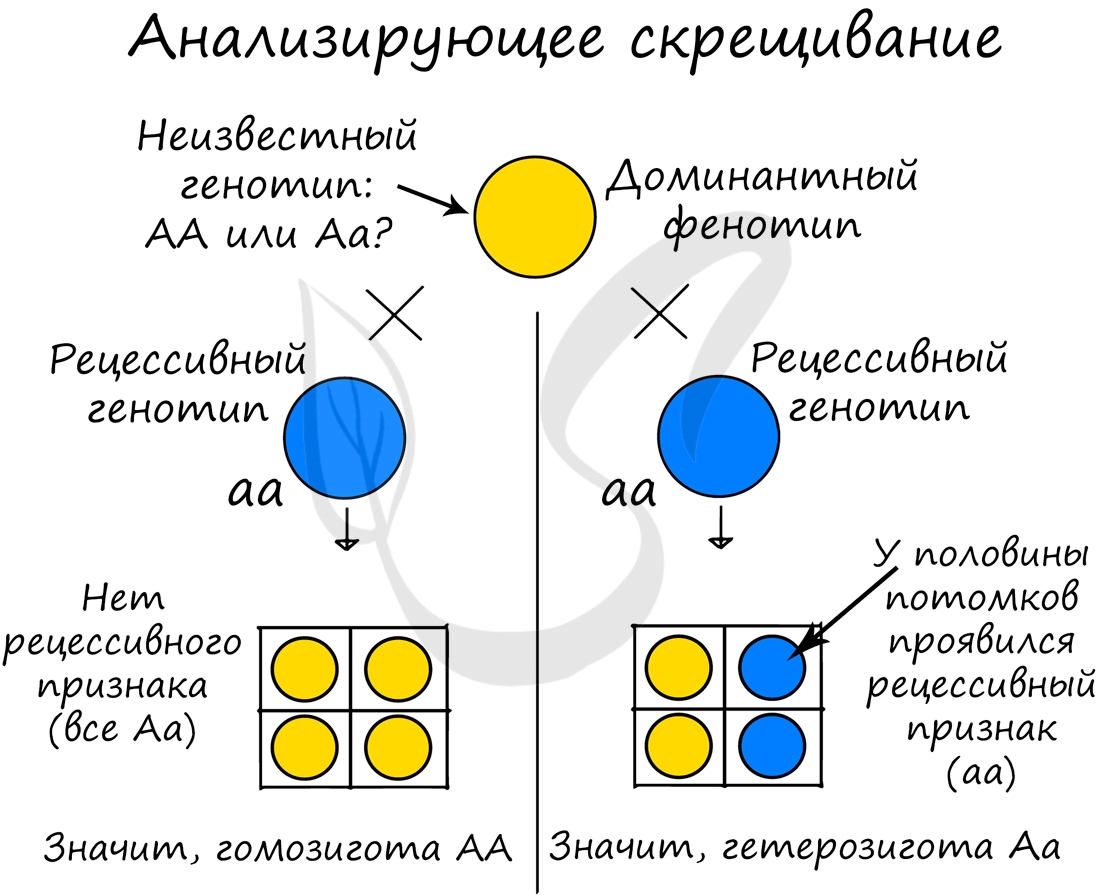
В рассмотренном случае, если генотип изучаемой особи содержит два доминантных гена (AA) — то в потомстве не может проявиться рецессивного признака, так как все потомство будет единообразно (Aa). Если изучаемая особь содержит рецессивный ген (Aа), то половина потомства будет его иметь (aa). В результате становится известен генотип гибридной особи.
Неполное доминирование
Помимо полного доминирования, существует неполное доминирование, которое характерно для некоторых генов. Известным примером неполного доминирования является наследование окраски лепестков у растения ночная красавица. В этом случае гены не полностью подавляют друг друга — проявляется промежуточный признак.
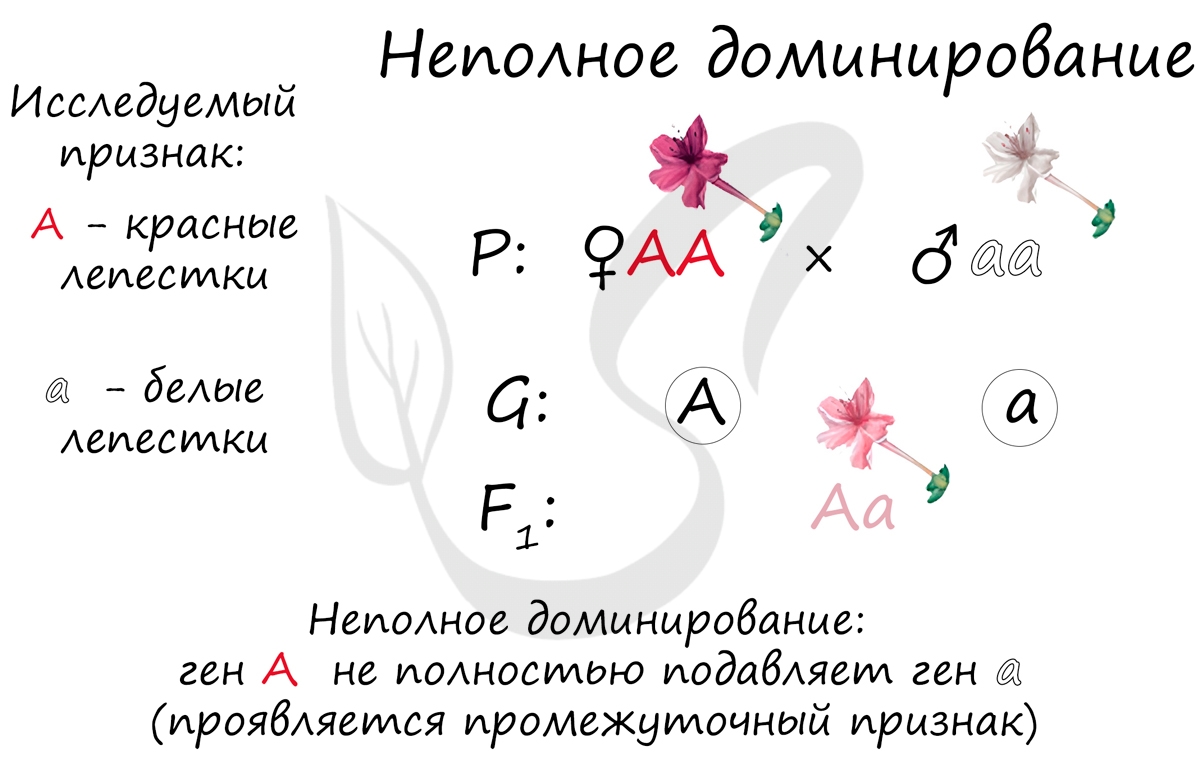
Обратите внимание, потомство F1 получилось также единообразным (возможен только один вариант — Aa), но фенотипически у гетерозиготы признак будет проявляться как промежуточное состояние (AA — красный, aa — белый, Aa — розовый). Это можно сравнить с палитрой художника: представьте, как смешиваются красный и белый цвета — получается розовый.
Второй закон Менделя — закон расщепления
«При скрещивании гетерозиготных гибридов (Aa) первого поколения F1 во втором поколении F2 наблюдается расщепление по данному признаку: по генотипу 1 : 2 : 1, по фенотипу 3 : 1″
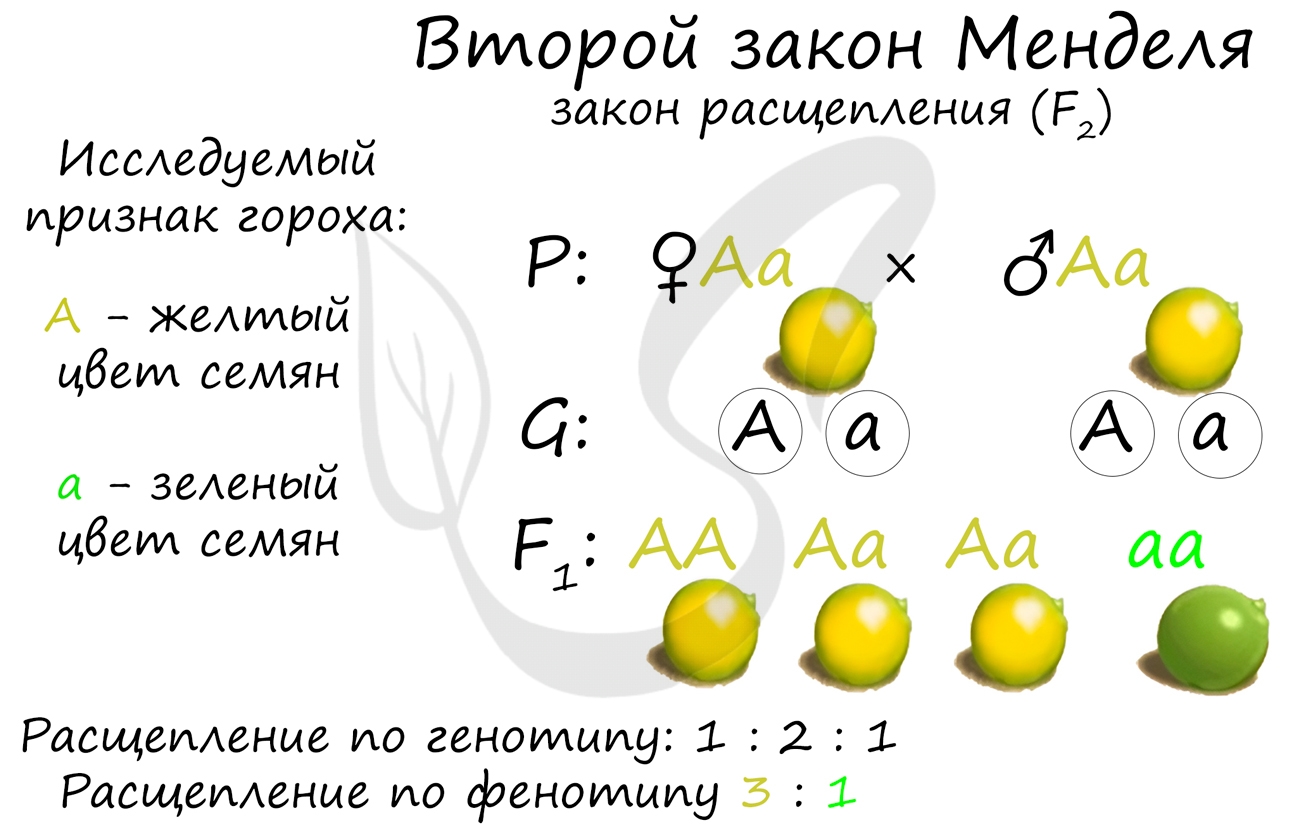
Скрещивая между собой гибриды первого поколения (Aa) Мендель обнаружил, что в потомстве особей с доминантным признаком (AA, Aa — желтый цвет семян) примерно в 3 раза больше, чем особей с рецессивным (aa).
Искренне желаю того, чтобы вы научились сами определять расщепление по генотипу и фенотипу. Это сделать не сложно: когда речь идет о генотипе, обращайте внимание только на гены (буквы), то есть если перед вами особи AA, Aa, Aa, aa — следует брать генотипы по очереди и складывать количество одинаковых генотипов. Именно в результате таких действий соотношение по генотипу получается 1:2:1.
Если перед вами стоит задача посчитать соотношение по фенотипу, то вообще не смотрите на гены — это только запутает! Следует учитывать лишь проявление признака. В потомстве получилось 3 растения с желтым цветом семян и 1 с зеленым, следовательно, расщепление по фенотипу 3:1.
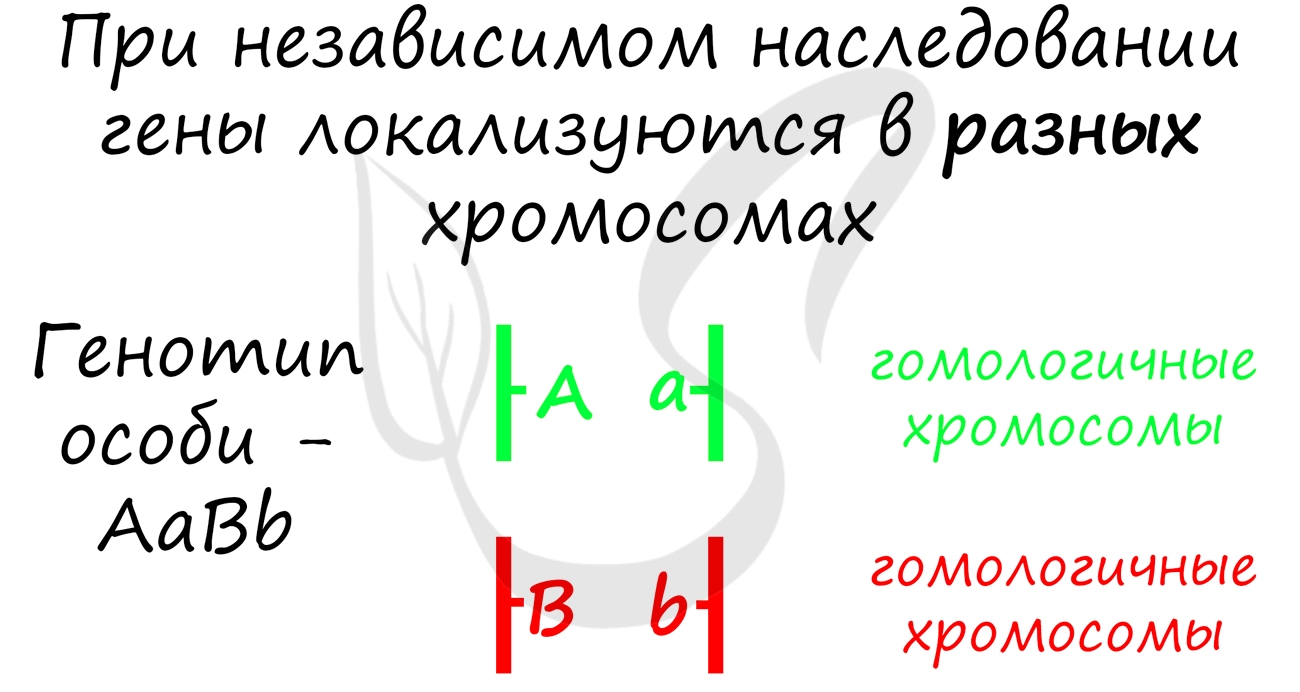
Третий закон Менделя — закон независимого наследования
В нем речь идет о дигибридном скрещивании, то есть мы исследуем не один, а два признака у особей (к примеру, цвет семян и форма семян). Каждый ген имеет два аллеля, поэтому пусть вас не удивляют генотипы AaBb 🙂 Важно заметить, что речь в данном законе идет о генах, которые расположены в разных хромосомах.
Запомните III закон Менделя так: «При скрещивании особей, отличающихся друг от друга по двум (и более) парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга, комбинируясь друг с другом во всех возможных сочетаниях.
Комбинации генов отражаются в образовании гамет. В соответствии с правилом, изложенным выше, дигетерозигота AaBb образует 4 типа гамет: AB, ab, Ab, aB. Повторюсь — это только если гены находятся в разных хромосомах. Если они находятся в одной, как при сцепленном наследовании, то все протекает по-другому, но это уже предмет изучения следующей статьи.
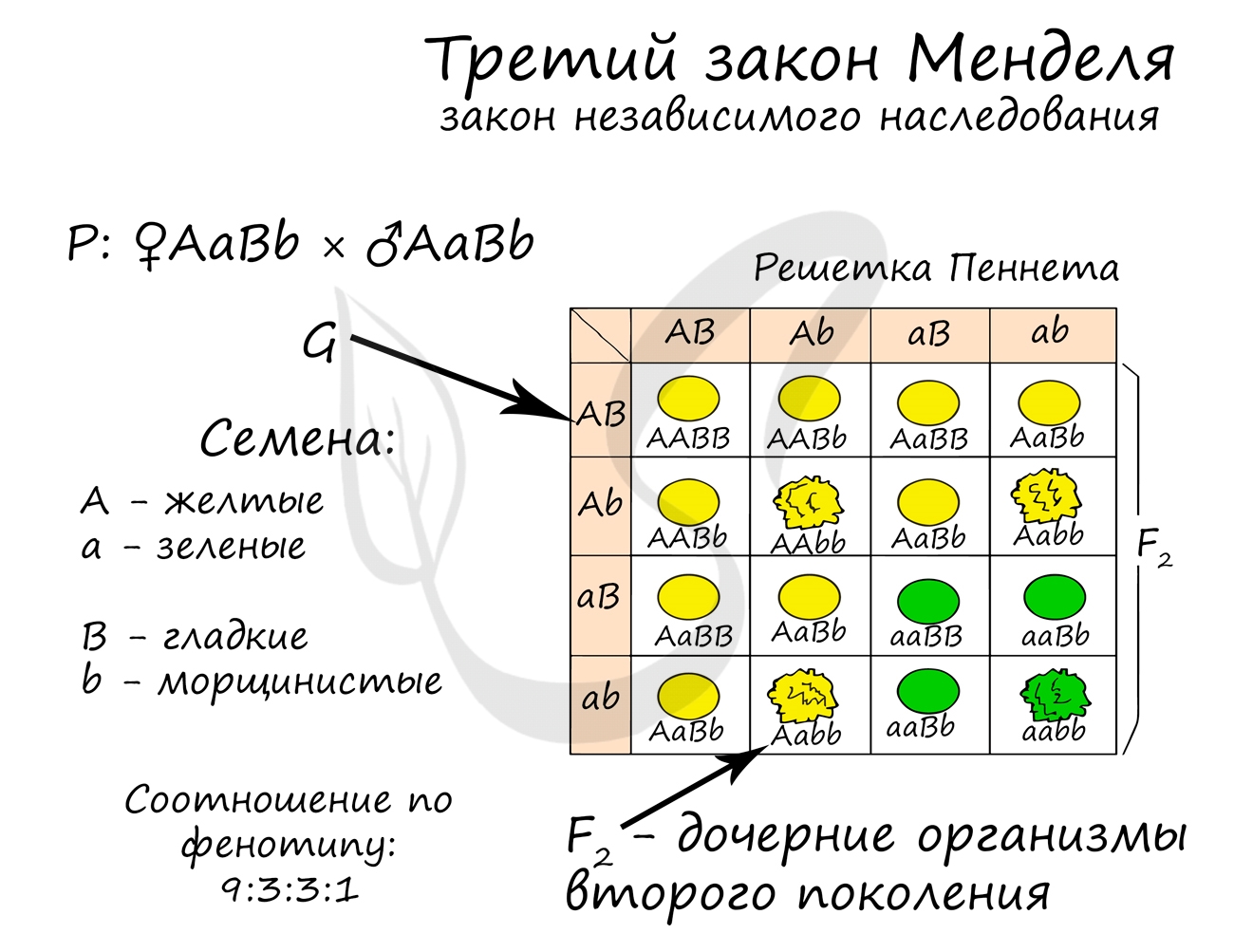
Каждая особь AaBb образует 4 типа гамет, возможных гибридов второго поколения получается 16. При таком обилии гамет и большом количестве потомков, разумнее использовать решетку Пеннета, в которой вдоль одной стороны квадрата расположены мужские гаметы, а вдоль другой — женские. Это помогает более наглядно представить генотипы, получающиеся в результате скрещивания.
В результате скрещивания дигетерозигот среди 16 потомков получается 4 возможных фенотипа:
- Желтые гладкие — 9
- Желтые морщинистые — 3
- Зеленые гладкие — 3
- Зеленые морщинистые — 1
Очевидно, что расщепление по фенотипу среди гибридов второго поколения составляет: 9:3:3:1.
Пример решения генетической задачи №1
Доминантный ген отвечает за развитие у человека нормальных глазных яблок. Рецессивный ген приводит к почти полному отсутствию глазных яблок (анофтальмия). Гетерозиготы имеют глазное яблоко малых размеров (микрофтальмия). Какое строение глазных яблок будет характерно для потомства, если оба родителя страдают микрофтальмией?
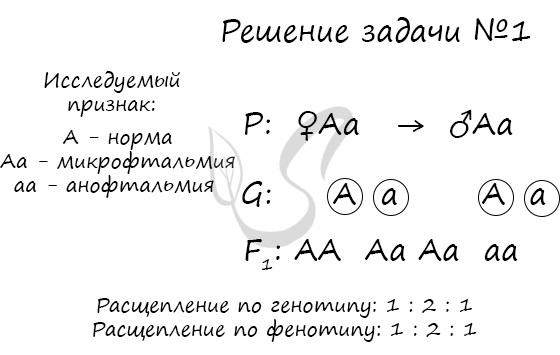
Обратите внимание на то, что доминирование генов неполное: человек с генотипом Aa будет иметь промежуточное значение признака — микрофтальмию. Поскольку доминирование неполное, то расщепление по генотипу и фенотипу совпадает, что типично для неполного доминирования.
В данной задаче только ¼ потомства (25%) будет иметь нормальные глазные яблоки. ½ часть потомства (50%) будет иметь глазное яблоко малых размеров — микрофтальмию, и оставшаяся ¼ (25%) будут слепыми с почти полным отсутствием глазных яблок (анофтальмией).
Не забывайте, что генетика, по сути, теория вероятности. Очевидно, что в жизни в такой семье может быть рождено 4 подряд здоровых ребенка с нормальными глазными яблоками, или же наоборот — 4 слепых ребенка. Может быть как угодно, но мы с вами должны научиться говорить о «наибольшей вероятности», в соответствии с которой с вероятностью 50% в этой семье будет рожден ребенок с микрофтальмией.
Пример решения генетической задачи №2
Полидактилия и отсутствие малых коренных зубов передаются как аутосомно-доминантные признаки. Гены, отвечающие за развитие этих признаков, расположены в разных парах гомологичных хромосом. Какова вероятность рождения детей без аномалий в семье, где оба родителя страдают обеими болезнями и гетерозиготны по этим парам генов.
Я хочу сразу навести вас на мысль о III законе Менделя (закон независимого наследования), который скрыт в фразе » Гены … расположены в разных парах гомологичных хромосом». Вы увидите в дальнейшем, насколько ценна эта информация. Также заметьте, что речь в этой задаче идет о аутосомных генах (расположенных вне половых хромосом). Аутосомно-доминантный тип наследования означает, что болезнь проявляется, если ген в доминантном состоянии: AA, Aa — болен.
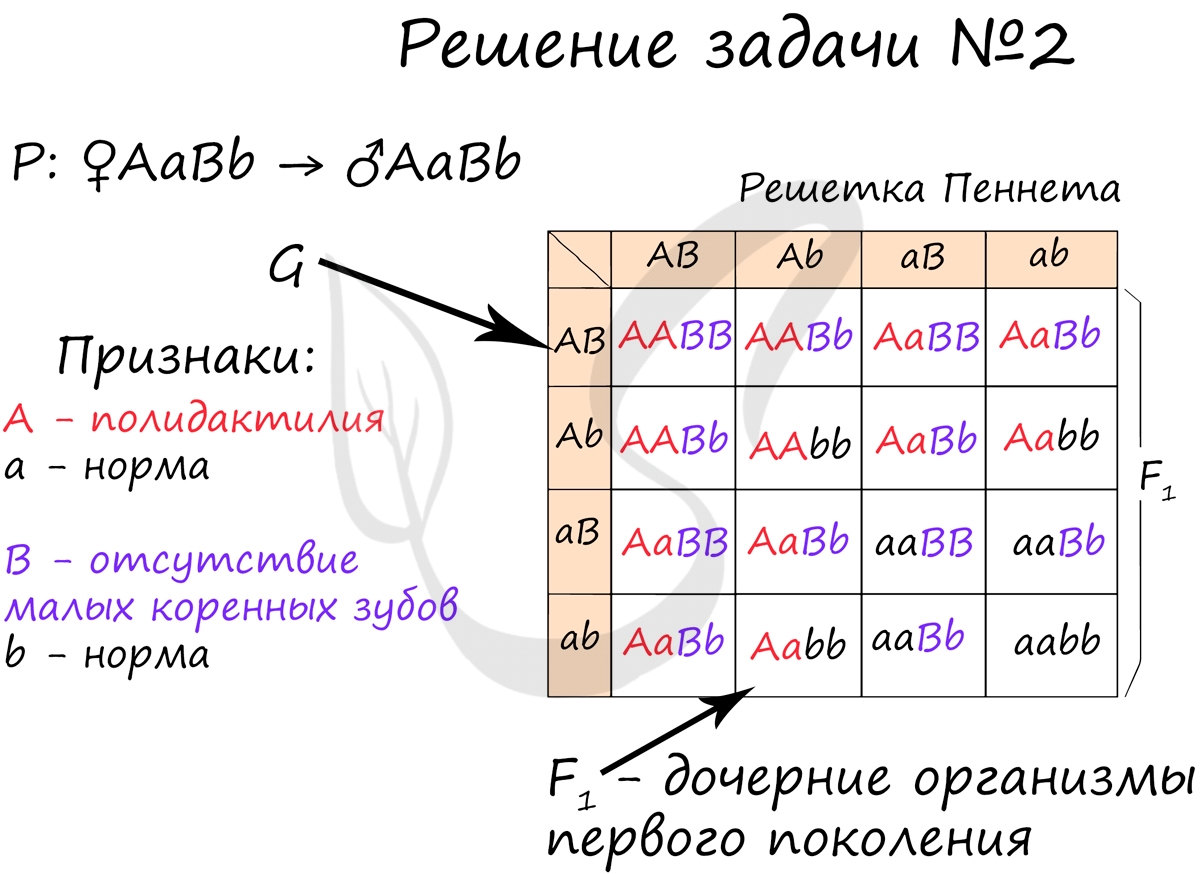
В данном случае мы построим решетку Пеннета, которая сделает генотипы потомства более наглядными. Вы видите, что на потомстве буквально нет ни одного живого места: почти все 16 возможных потомков больны либо одним, либо другим заболеванием, кроме одного, aabb. Вероятность рождения такого ребенка очень небольшая 1/16 = 6.25%.
Пример решения генетической задачи №3
У голубоглазой близорукой женщины от брака с кареглазым мужчиной с нормальным зрением родилась кареглазая близорукая девочка и голубоглазый мальчик с нормальным зрением. Ген близорукости (A) доминантен по отношению к гену нормального зрения (a), а ген кареглазости (D) доминирует над геном голубоглазости (d). Какова вероятность рождения в этой семье нормального кареглазого ребенка?

Первый этап решения задачи очень важен. Мы учли описания генотипов родителей и, тем не менее, белые пятна остались. Мы не знаем гетерозиготна (Aa) или гомозиготная (aa) женщина по гену близорукости. Такая же ситуация и с мужчиной, мы не можем точно сказать, гомозиготен (DD) он или гетерозиготен (Dd) по гену кареглазости.
Разрешение наших сомнений лежит в генотипе потомка, про которого нам рассказали: «голубоглазый мальчик с нормальным зрением» с генотипом aadd. Одну хромосому ребенок всегда получает от матери, а другу от отца. Выходит, что такого генотипа не могло бы сформироваться, если бы не было гена a — от матери, и гена d — от отца. Следовательно, отец и мать гетерозиготны.

Теперь мы можем точно сказать, что вероятность рождения в этой семье нормального кареглазого ребенка составляет ¼ или 25%, его генотип — Ddaa.
Аутосомно-доминантный тип наследования
Я не забыл о том, что по ходу изучения генетики вас надо научить видеть различные варианты наследования на генеалогическом древе (родословной) =) Из предыдущей статьи мы узнали о том, как выглядит и чем характеризуется аутосомно-рецессивный тип наследования, сейчас поговорим об аутосомно-доминантном, с которым мы столкнулись в задачах выше.
Аутосомно-доминантный тип наследования можно узнать по следующим признакам:
- Болезнь проявляется в каждом поколении семьи (передача по вертикали)
- Здоровые дети больных родителей имеют здоровых детей
- Мальчики и девочки болеют одинаково часто
- Соотношение больных и здоровых 1:1
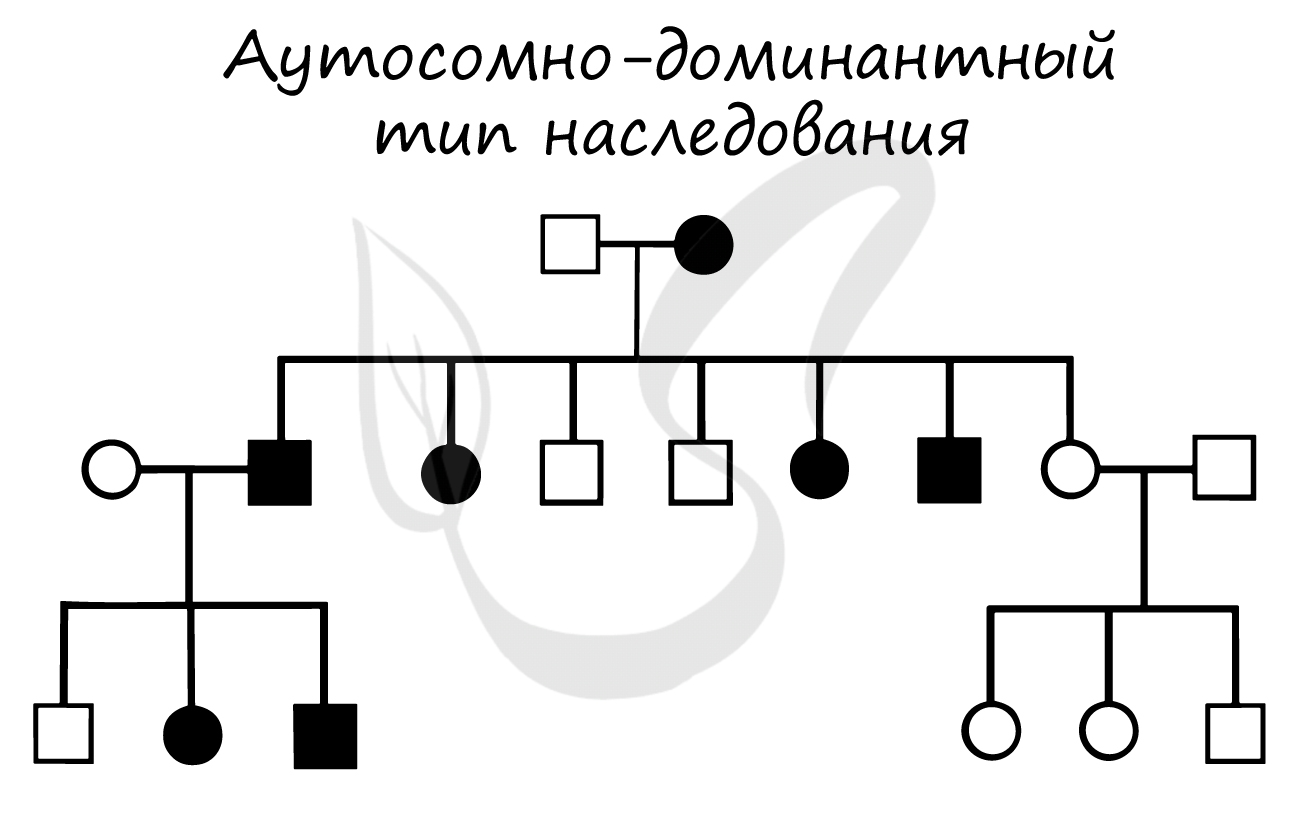
©Беллевич Юрий Сергеевич
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Закони Менделя — Вікіпедія

Зако́ни Ме́нделя — закони, що становлять основу класичної генетики. У своїх працях Грегор Мендель ґрунтувався на дослідженнях, проведених на горосі посівному (рід Pisum). Цей об’єкт виявився вдалим, тому що для нього характерне самозапилення, яке уможливлює одержання чистих ліній, тобто особин гомозиготних за більшістю генів. У своїх роботах Мендель не виділяв окремих законів, їх виділили й назвали інші дослідники, вже після їхнього перевідкриття в 1900 році.
Уявлення про спадковість до робіт Менделя[ред. | ред. код]
Навіть у найдавніших писемних пам’ятках, що дійшли до наших днів, згадуються деякі особливості спадковості, такі як схожість між членами однієї сім’ї. Основні уявлення про спадковість, поширені до початку XX століття, базувались на двох принципах. По-перше, вважалось, що спадковість працює тільки в межах одного виду. Хоч у міфології багатьох народів стрічаються такі гібридні тварини як мінотаври, кентаври, мантикори тощо, а жирафи вважались наслідком схрещення верблюда і леопарда (це відображено у видовій назві Giraffa camelopardalis), починаючи із Середньовіччя людям стало відомо, що гібридизація між такими далекими видами переважно неможлива. Тому вважалося, що спадковість і мінливість діє тільки всередині кожного виду, а самі види залишились незмінними від часу їхнього створення.
Інше «доменделівське» уявлення про спадковість полягало у тому, що риси успадковуються безпосередньо. Так Гіппократ писав, що репродуктивний матеріал, який він називав «ґонос», від кожної частини тіла батьків незалежно передається відповідній частині тіла дитини і визначає її розвиток. Схожі теорії спадковості були поширені ще до кінця XIX ст. Наприклад, Дарвін 1868 року припускав, що тканини тіла батьків продукують мікроскопічні гранули (gemmules), які передаються дитині і забезпечують формування відповідних структур в ембріона. Уявлення про пряме успадкування переважно поєднувалось із ідеєю про змішування ознак матері і батька. Наприклад, якщо в одного з батьків чорне волосся, а в іншого — біле, діти повинні бути русявими.
Суперечливість «класичних» уявлень[ред. | ред. код]
 Йозеф Готліб Кельрейтер
Йозеф Готліб Кельрейтер«Класичні» уявлення про спадковість, проте, породжували очевидний парадокс: якщо, по-перше, спадковість існує тільки в межах одного виду і немає жодних зовнішніх джерел мінливості, по-друге, риси батьків просто змішуються у потомства, тоді з часом всі особини одного виду мали би стати зовсім однаковими. А це аж ніяк не спостерігається у природі.
Перші наукові результати, що ставили під сумнів ідею про пряме успадкування, отримав 1760 року німецький ботанік Йозеф Готліб Кельрейтер. Він схрещував різні сорти тютюну і спостеріг, що перше покоління нащадків відрізнялось від батьківських особин. Проте, якщо схрестити ці рослини між собою, то в наступному поколінні серед усієї різноманітності отриманих рослин можна знайти не тільки дуже схожих до особин першого покоління (своїх батьків), а й до вихідних сортів (своїх «дідусів і бабусь»). Ця робота стала першим кроком до розвитку сучасної генетики. Кельрейтер чітко показав, що, по-перше, ознаки не змішуються, а, по-друге, можуть «маскуватись» в одному поколінні і знову виникати в наступному. Це останнє спостереження очевидно суперечило уявленню про пряме успадкування.
Впродовж наступної сотні років робота у напрямку, започаткованому Кельрейтером, продовжувалась. Зокрема, над вдосконаленням культурних рослин працювали деякі англійські джентельмени. Один із них, Т.А. Найт, у 1790-их роках схрестив дві чисті лінії горошку (Pisum sativum) із білими і рожевими квітами. Усі рослини першого покоління мали рожеві квіти, проте у другому знову з’явилась невелика кількість особин із білими пелюстками. Отже результати Найта підтверджували більш ранні висновки Кельрейтера. Проте, ці роботи мали один вагомий недолік: вони не містили конкретних числових даних. Кельрейтер у своїх пізніх працях писав, що деякі ознаки мають «сильнішу тенденцію» проявлятися, але не пробував описати цю тенденцію математично. Це стало однією із причин того, що «революція» у розумінні спадковості затрималась на століття. Ще одну важливу працю, яка, ймовірно, вплинула на формування ідей Менделя, опублікував у 1863 Чарльз Ноден. У ній французький натураліст обговорює в загальному експерименти із гібридизації рослин, а також повідомляє про розщеплення ознак і домінантність (термін введений Сажере 1826 року) у дурману. Проте Ноден також не описує свої результати математично[2]. Саме базуючись на ідеях попередників, і поєднавши їх із вдалим плануванням експериментів і використанням математичного апарату, Менделю вдалось відкрити основні принципи спадковості.
 Монастир у Брно, де працював Мендель
Монастир у Брно, де працював МендельГрегор Мендель народився 1822 року у Моравії. По закінченню школи і Оломоуцького інституту філософії у віці 21 рік він постригся в монахи Августинського монастиря Святого Томаса у місті Брюнн (зараз Брно, Чехія). У той час в Австро-Угорській імперії політика стосовно релігійних закладів, полягала у тому, що вони повинні були служити не тільки Церкві, а й державі. Через це, монахи мали доступ до великих бібліотек, колекцій гербаріїв, і загалом вели багате інтелектуальне життя[4].
Економічні фактори, такі як розвиток торговельної мережі у Європі, стимулювали розвиток досліджень у галузі сільського господарства. У 1806 році Крістіан Карл Андре заснував Моравське товариство сільського господарства (повна назва: Моравське товариство удосконалення сільського господарства, природничих наук і знань про село). Абат Монастиря Святого Томаса Непп засідав у комітеті цього товариства, а також був президентом Асоціації Помологів і Енологів. Мендель був зацікавлений у вивчені природничих наук і сільського господарства, і абат монастиря його підтримував у цьому. Він посприяв вступу Грегора у Віденський університет, де він вивчав природничі науки і математику, зокрема відвідував курси фізіології рослин Франца Унгера і експериментальної фізики Крістіана Допплера[4]. Не склавши випускних іспитів в університеті Мендель повернувся до монастиря, де провів решту свого життя, і взявся за дослідження гібридизації рослин. Тут у садку монастиря починаючи із 1857 року він проводив досліди, що заклали основи сучасної генетики. Свої результати Мендель представив 1865 року на засіданні Брюннського товариства природознавства, а в 1866 році опублікував їх у записках цієї організації.
Вибір об’єкта[ред. | ред. код]

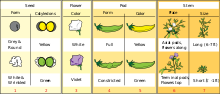 Сім ознак, які Мендель вибрав для аналізу
Сім ознак, які Мендель вибрав для аналізуЯк модель для своїх досліджень Грегор Мендель вибрав горох посівний (Pisum sativum), той же об’єкт, на якому проводили експерименти Найт та багато інших дослідників. Для цього було кілька причин. По-перше, попередні дані, отримані у працях із цією рослиною, свідчили про те, що можна очікувати розщеплення ознак у потомства. По-друге, у торговців насінням можна було купити різні сорти цієї рослини, що відрізнялись між собою чіткими ознаками, такими як забарвлення віночка або насіння. Мендель почав аналіз і ретельного вивчення 32-ох чистих ліній горошку, і вибрав ті, що відрізнялись за сімома ознаками.
По-третє, горошок здатний як до самозапилення так і до перехресного запилення. Через те, що тичинки і маточка у квітах цієї рослини оточені двома зрослими пелюстками (так званий човен), то пилок із перших переноситься на приймочку останньої, так відбувається самозапилення. Якщо ж дослідник хоче здійснити перехресне запилення, то вирізає тичинки у квітах «материнської» рослини до того, як вони дозріють, і переносить пилок із «батьківської» рослини пензликом або безпосередньо тичинками.
Окрім того, горошок вигідний із практичних міркувань: він дешевий, невибагливий, не займає багато місця, має відносно короткий цикл розвитку і дає достатньо багато потомства. Тому можна аналізувати одночасну велику кількість рослин і вирощувати кілька поколінь впродовж одного року, що прискорює отримання результатів.
Особливості підходу[ред. | ред. код]
Причиною успіху дослідів Менделя порівняно із його попередниками було вдале планування експериментів. По-перше, він аналізував тільки сім ознак, що мають два чітко відмінні один від одного стани, такі як гладкість/зморшкуватість насіння, зелене/жовте забарвлення сім’ядоль, фіолетові/білі квіти (і, відповідно, сіра/біла шкірка насіння), повні/стягнуті боби, зелене/жовте забарвлення бобів, осьове/термінальне розташування квітів і плодів, високий ріст (180—215 см)/карликовість (20—30 см). Якби Мендель натомість віддав перевагу дослідженню ознак із неперервним розподілом, таких, як, наприклад, маса насінин, то не зміг би відкрити дискретної природи спадковості. По-друге, він почав свою роботу із того, що перевірив, чи вибрані сорти є справді чистими лініями, тобто чи кожне покоління, отримане від самозапилення, буде зберігати всі батьківські ознаки. На виконання цієї перевірки він затратив два роки. По-третє, він вибрав математичний підхід до опису результатів і обробляв їх статистично.
Постановка типового досліду[ред. | ред. код]
Після того, як Мендель пересвідчився, що обрані сорти є чистими лініями, він проводив експерименти із гібридизації. Перші схрещування були моногібридними, тобто такими, у яких враховувалась якась тільки одна ознака, наприклад, колір квітів. Мендель проводив обернені схрещування, тобто, якщо у прямому він переносив пилок із рослин з білими квітами на приймочки рослин із фіолетовими квітами, то в оберненому навпаки. Це робилось для того, щоб пересвідчитись, що походження статевих клітин не впливає на прояв ознаки у нащадків. У випадку тих рис, які аналізував він, обернені схрещування справді давали однакові результати.
Після отримання гібридів першого покоління (F1) Мендель дозволяв їм самозапилитись, для того, щоб відбулось розщеплення альтернативних станів ознак у другому поколінні (F2). Після цього він рахував всі рослини із конкретним фенотипом. У деяких дослідах він простежував успадкування ознак до третього покоління.
Аналогічним чином Мендель проводив також і ди- і тригібридні схрещування, у яких батьківські особини відрізнялись за двома чи трьома ознаками відповідно, щоб з’ясувати, як успадкування однієї із них впливає на успадкування іншої. Нарешті, для підтвердження результатів, отриманих у дослідах на горошку, він також проводив схожі експерименти із квасолею (Phaseolus vulgaris і Phaseolus nanus)[13].
Закономірності спадковості, встановлені Менделем[ред. | ред. код]
Моногібридні схрещування[ред. | ред. код]
Перший закон[ред. | ред. код]
Схрещуючи рослини гороху, що відрізнялись за станами однієї ознаки, Мендель спостеріг, що у фенотипі всіх гібридів першого покоління (F1) проявлявся тільки один із двох станів. Наприклад, всі рослини, отримані внаслідок гібридизації між горохом із білими квітами і горохом із фіолетовими квітами, мали фіолетові квіти. Такі результати підтверджували роботи Найта та інших попередників Менделя і заперечували уявлення про «змішування» ознак батьків у потомства.
Стан ознаки, який проявлявся в F1, Мендель називав домінантним, а той, який не проявлявся —рецесивним. Для всіх семи пар станів ознак, які аналізував Мендель, один виявився домінантним, інший — рецесивним.
| Перший закон Менделя, або закон одноманітності гібридів першого покоління, формулюється так: |
| У першому поколінні від схрещування гомозигот із домінантною та рецесивною ознаками виявляється тільки домінантна ознака. |
Другий закон[ред. | ред. код]
Після самозапилення особин F1 Мендель зібрав і висадив насіння із кожної рослини, щоб проаналізувати друге покоління. Цього разу серед рослин з’явились такі, що несли рецесивну ознаку (тобто ту, яка зовсім не виникала в F1). Щоб краще зрозуміти, яким чином відбувається успадкування, Мендель порахував всі особини, у яких проявлялась певна ознака. Наприклад, при схрещуванні гороху із білими і фіолетовими квітами, в F2 було всього 929 рослини, із яких 705 мали фіолетові квіти, і 224 — білі. Результати всіх схрещувань наведені у таблиці.
| Фенотип батьків | F1 | F2 | Співвідношення фенотипів у F2 |
|---|---|---|---|
| Кругле × зморшкувате насіння | Всі з круглим насінням | 5474 круглі; 1850 зморшкуваті | 2.96 : 1 |
| Зелене × жовте насіння | Усі із жовтим насінням | 6022 жовті; 2001 зелені | 3.01 : 1 |
| Фіолетові × білі квіти | Усі із фіолетовими квітами | 705 фіолетові; 224 білі | 3.15 : 1 |
| Стягнуті × повні боби | Усі з повними бобами | 882 повні; 299 стягнуті | 2.95 : 1 |
| Зелені × жовті боби | Усі із зеленими бобами | 428 зелені; 152 жовті | 2.82 : 1 |
| Верхівкове × осьове розташування квітів | Усі із осьовими квітами | 651 осьові; 207 верхівкові | 3.14 : 1 |
| Високі × низькі стебла | Усі із високими стеблами | 787 високі; 277 низькі | 2.84 : 1 |
Таким чином в середньому співвідношення фенотипових класів у другому поколінні становило 3:1, тобто у четвертини особин проявлялась рецесивна ознака.
| Другий закон Менделя, або закон розщеплення, говорить: |
| При схрещуванні гібридів першого покоління у нащадків спостерігається розщеплення фенотипових класів у співвідношенні 3:1. |
Після цього Мендель взявся за аналіз особин F2, які проявляли домінантну ознаку, і виявив, що насправді вони не є однорідним класом, а відрізняються одне від одного генетично. У цьому випадку він працював із рослинами, що мали різне забарвлення насіння (жовте — домінантна ознака, зелене — рецесивна). Вибір саме цієї ознаки був зручним через те, що колір кожної горошини визначається її власним генотипом, а не генотипом материнської особини. Тому насінину можна трактувати як окрему особину і оцінювати її фенотип без потреби пророщувати, як це необхідно у випадку багатьох інших ознак. Після схрещування чистих ліній із жовтим і зеленим насінням за описаною вище схемою, Мендель відібрав 519 жовтих горошин F2, проростив їх і дозволив самозапилення. Після цього із кожної рослини окремо зібрав насіння. З’ясувалось, що у 166 особин усе насіння було жовтим, а в решти 353 спостерігалось розщеплення 3:1 між класами із жовтим і зеленим насіння.
Провівши аналогічний аналіз особин F2 із зеленими сім’ядолями Мендель пересвідчився, що на відміну від рослин із домінантною ознакою, ці були чистою лінією. Таким чином стало зрозуміло, що за фенотиповим співвідношенням 3:1 криється більш фундаментальне генотипове співвідношення 1:2:1. Це підтвердилось і для інших ознак, які аналізував Менедель. Отже, його модель спадковості пояснювала не розщеплення у другому поколінні 3:1, а саме 1:2:1. Ця модель є прикладом вдалої побудови на основі експериментальних даних наукової гіпотези, що підлягає подальшому дослідному тестуванню.
| Третій закон Менделя, або закон незалежного успадкування, говорить: |
| При схрещуванні двох особин, що відрізняються один від одного за двома (і більше) парам альтернативних ознак, гени і Відповідні їм ознаки успадковуються незалежно один від одного і комбінуються у всіх можливих поєднаннях (як і при моногібридному схрещуванні ). |
Коли схрещувалися гомозиготні рослини, що відрізняються за кількома ознаками, такими як білі і фіолетові квіти і жовте або зелене насіння, успадкування кожної з ознак відповідало першим двом законам і в потомстві вони комбінувалися таким чином, ніби їхнє успадкування відбувалося незалежно одне від одного. Перше покоління після схрещування мало домінантний фенотип за всіма ознаками. У другому поколінні спостерігалося розщеплення фенотипів за формулою 9: 3: 3: 1, тобто 9:16 були з фіолетовими квітами і жовтим насінням, 3:16 з білими квітами і жовтим насінням, 3:16 з фіолетовими квітами і зеленим насінням, 1 : 16 з білими квітами і зеленим насінням.
Менделю трапилися ознаки, гени яких знаходилися в різних парах гомологічних хромосом (нуклеопротеїдних структур в ядрі еукаріотичної клітини, в яких зосереджена велика частина спадкової інформації і які призначені для її зберігання, реалізації і передачі) гороху. При мейозі гомологічні хромосоми різних пар комбінуються в гаметах випадковим чином. Якщо в гамету потрапила батьківська хромосома першої пари, то з однаковою ймовірністю в цю гамету може потрапити як батьківська, так і материнська хромосома другої пари. Тому ознаки, гени яких знаходяться в різних парах гомологічних хромосом, комбінуються незалежно одна від одної. (Згодом з’ясувалося, що з досліджених Менделем семи пар ознак у гороху, у якого диплоїдне число хромосом 2n = 14, гени, що відповідають за одну з пар ознак, перебували в одній і тій же хромосомі. Однак Мендель не виявив порушення закону незалежного успадкування, оскільки зчеплення між цими генами не спостерігалося через велику відстань між ними).
Менделева модель спадковості і її пояснення[ред. | ред. код]
Щоб пояснити отримані результати, Мендель запропонував модель спадковості, що складається із кількох припущень, які в результаті лягли в основу класичної генетики.
По-перше, ознаки не передаються безпосередньо від батьків нащадкам, натомість діти успадковують певні дискретні частинки, які несуть інформацію про конкретні ознаки. Пізніше ці часточки спадковості беруть участь у формуванні відповідних рис. Мендель називав їх «факторами», а в сучасній генетиці вони позначаються терміном «ген».
По-друге, кожна особина має по дві копії кожного виду цих часточок спадковості, тобто генів. Вони можуть бути однаковими або відрізнятись. Варіанти гену, які визначають різні стани однієї ознаки, згідно із сучасною термінологією називаються алелями, організми, які несуть два однакові алелі певного гену називають гомозиготними за цим геном, а ті, що несуть різні алелі, — гетерозиготними.
Два алелі одного гену, наявні в однієї особини, не впливають одне на одного, вони не можуть зливатись між собою чи модифікувати одне одного. Через, що залишаються, як висловлюється Мендель, «незабрудненими». Під час утворення статевих клітин (гамет), кожна із них отримує тільки одну із двох наявних у соматичних клітинах «чисту» копію кожного гену. При чому розподіл відбувається порівну: тобто 50% гамет організму гетерозиготного за певним геном нестимуть один алель, а 50% — інший. Цей принцип ще називають правилом (законом) чистоти гамет.
| Правило чистоти гамет: |
| Кожна гамета диплоїдного гетерозиготного організму несе лише один алельний ген і не може одночасно містити обидва. |
Запліднення відбувається випадковим чином, тобто гамети комбінуються між собою не в залежності від того, які саме алелі вони несуть. І нарешті, наявність в організму певного алелю ще не гарантує, що він проявиться у фенотипі, у гетерозиготних організмів розвивається тільки один із двох альтернативних станів ознаки, який називають домінантним.
Перевірка моделі[ред. | ред. код]
Менделева модель спадковості чудово пояснювала його експериментальні дані. Проте щоб перевірити її справедливість, необхідно було зробити теоретичні передбачення на основі своєї гіпотези, після чого пересвідчитись, що вони відповідатимуть реальності. Для цього він використав аналізуючі схрещування. Одним із ключових моментів у запропонованій моделі було те, що особини F1 несуть два різні алелі (тобто є гетерозиготами), які рівномірно розділяються між гаметами. Отже, якщо схрестити ці рослини із рослинами гомозиготними за рецесивним алелем, то у наступному поколінні повинне виникнути розщеплення за фенотипом 1:1. Провівши гібридизацію особин F1 із жовтими сім’ядолями, отриманих від схрещування чистих ліній із зеленими і жовтими сім’ядолями, із рослинами з рецесивною ознакою (зеленими сім’ядолями), Мендель отримав розщеплення близьке до 1:1 (58 жовті і 52 зелені).
Клітинні і молекулярні основи менделівського успадкування[ред. | ред. код]
Фізичні носії спадковості[ред. | ред. код]
- Див. також: Ген, ДНК
Оригінальна стаття Менделя містила багато абстракцій, таких як «фактори спадковості». Крім того, на сучасному йому етапі розвитку біологічної науки не було відомо багатьох процесів і явищ, які дозволили б пояснити механізми домінантності певних алелів, утворення «чистих» гамет тощо. Проте накопичені з того часу знання дозволили підтвердити і доповнити Менделеву модель. Так стало відомо, що «фактори спадковості» або, згідно із сучасною термінологією, гени — це ділянки ДНК, у яких найчастіше закодована інформація, потрібна для синтезу певного поліпептиду (хоча бувають інші типи генів). Різні алелі одного гену відрізняються послідовністю нуклеотидів у них, хоч, зазвичай незначно. Так, на кілька тисяч пар нуклеотидів зміненою переважно є тільки одна або кілька позицій. Для певного гену може існувати велика кількість варіантів, тобто алелів.
Домінантність ознак[ред. | ред. код]
- Див. також: Домінантність (генетика)
Алелі виникають внаслідок мутацій і їх подальшого закріплення у популяції. Ці мутації часто призводять до того, що продукт гену повністю втрачає свою функціональність. Наприклад, для утворення фіолетового забарвлення пелюсток горошку необхідний пігмент групи антоціанів. Ця хімічна речовина синтезується клітинами у ряді реакцій, що каталізуються ферментами. Останні є білками, інформація про структуру яких закодована у відповідних генах. Рецесивна ознака — біле забарвлення віночка — виникає тоді, коли немає одного із ферментів цього біохімічного шляху. І проявляється вона тільки в тому разі, коли «зламані» обидві копії гену диплоїдного організму (генотип aa), оскільки одного функціонального алелю часто буває цілком достатньо для виконання потрібної роботи. Слід зазначити, що це тільки один із відомих механізмів домінування одних ознак над іншими.
Розщеплення ознак[ред. | ред. код]
- Див. також: Мейоз
Правило чистоти гамет і закон розщеплення вдалось пояснити після того, як стало зрозуміло, що хромосоми є носіями спадковості у клітинах, і відкриття мейозу. Соматичні клітини багатьох організмів, зокрема горошку і людей, є диплоїдними, тобто містять два набори хромосом, отримані від двох батьківських організмів під час запліднення. Кожна хромосома із пари несе один із двох алелів, при чому розташовані вони в однаковому місці (локусі). Утворенню статевих клітин передує спеціальний тип поділу ядра — мейоз. Під час цього поділу відбувається розділення наборів хромосом. Тобто із утворених внаслідок мейозу гаплоїдних клітин половина міститиме одну хромосому із пари, половина — іншу. Або, іншими словами, для гетерозиготи Aa половина гамет нестиме алель A, половина — a.
На основі інформації про те, що різні типи гамет утворюються у гетерозиготи із рівною імовірністю, а запліднення відбувається випадковим чином, можна оцінити імовірність отримання певного фенотипового класу у потомстві. Наприклад, при схрещуванні двох гетерозигот Aa і імовірність появи серед нащадків особин із рецесивною ознакою буде рівна ½ (ймовірність того, що яйцеклітина міститиме алель a) × ½ (ймовірність того, що сперматозоїд міститиме алель a) = 1/4, що відповідає розщепленню 1:3. Такі імовірності можна візуалізувати за допомогою простої діаграми, яка називається решіткою Пеннета. Це двомірна таблиця, у якій у клітинах по горизонталі записують всі можливі типи гамет одного батьківського організму, а по вертикалі — іншого. У клітинках на перетині зазначають можливі комбінації цих гамет, при чому кожна із цих комбінацій має однакову ймовірність виникнення.
Незалежне успадкування ознак[ред. | ред. код]
- Третій закон або «Закон незалежного успадкування ознак»: кожна пара альтернативних варіантів ознак успадковується незалежно від інших пар і дає розщеплення 3:1 за кожною з пар (як і при моногібридному схрещуванні). При дигібридному схрещуванні (коли спостереження ведеться за двома ознаками) серед гібридів другого покоління спостерігають розщеплення 9:3:3:1. Цей закон справедливий лише для ознак, у яких гени, що їх кодують, належать до різних груп зчеплення, тобто знаходяться в різних хромосомах. Закон може виконуватись і для ознак, гени яких знаходяться в одній хромосомі на значній відстані один від одного (не менше 50 морганід). В іншому випадку гени спадкуватимуться зчеплено.
- Менделю потрапили ознаки, гени яких знаходилися в різних парах гомологічних хромосом (нуклеопротеїдних структур в ядрі еукаріотичної клітини, в яких зосереджена велика частина спадкової інформації і які призначені для її зберігання, реалізації і передачі) гороху. При мейозі гомологічні хромосоми різних пар комбінуються в гаметах випадковим чином. Якщо в гамету потрапила батьківська хромосома першої пари, то з однаковою ймовірністю в цю гамету може потрапити як батьківська, так і материнська хромосома другої пари. Тому ознаки, гени яких знаходяться в різних парах гомологічних хромосом, комбінуються незалежно один від одного. (Згодом з’ясувалося, що з досліджених Менделем семи пар ознак у гороху, у якого диплоидное число хромосом 2n = 14, гени, що відповідають за одну з пар ознак, перебували в одній і тій же хромосомі. Однак Мендель ні виявив порушення закону незалежного успадкування, так як зчеплення між цими генами не спостерігалося через велику відстань між ними).
- Campbell NA, Reece JB (2008). Biology (вид. 8th). Benjamin Cammings. ISBN 978-0321543257.
- Griffiths A.J.F., Wessler S.R., et al (2004). An Introduction to Genetic Analysis (вид. 8th). W. H. Freeman. ISBN 978-0716749394. Архів оригіналу за 26 лютий 2015. Процитовано 13 серпень 2013.
- Raven P, Johnson G, Singer S, Losos J (2004). Biology (вид. 7th). McGraw-Hill. ISBN 978-0072921649.
- Sturtevant A.H. (2001 (original 1965)). A History of Genetics. Cold Spring Harbor Laboratory Press and Electronic Scholarly Publishing Project. ISBN 0-87969-607-9.
- Tamarin RH (2001). Principles of Genetics (вид. 7th). Mcgraw-Hill. ISBN 0072334193.
- Кучеренко М.Є., Вервес Ю.Г., Балан П.Г., Войціцький П.М. Загальна біологія. — 3-є. — Київ : Генеза, 2006. — ISBN 966-504-199-1.
- Сиволоб А.В., Рушковський С.Р., Кир’яченко С.С. та ін. Генетика: підручник. — Київ : ВПЦ «Київський університет», 2008. — ISBN 975-966-439-108-2.
Третий закон Менделя
☰
Третий закон Менделя — это закон независимого распределения признаков. Под этим подразумевается, что каждый ген одной аллельной пары может оказаться в гамете с любым другим геном из другой аллельной пары. Например, если организм гетерозиготен по двум исследуемым генам (AaBb), то он образует следующие типы гамет: AB, Ab, aB, ab. То есть, например, ген A может оказаться в одной гамете как с геном B, так и b. Это же касается и других генов (их произвольного сочетания с неаллельными генами).
Третий закон Менделя проявляется уже при дигибридном скрещивании (тем более при тригибридном и полигибридном), когда чистые линии различаются по двум исследуемым признакам. Мендель скрестил сорт гороха с желтыми гладкими семена с сортом, у которого были зеленые морщинистые семена, и получил исключительно желтые гладкие семена F1. Далее он вырастил из семян растения F1, позволил им самоопыляться и получил семена F2. И здесь он наблюдал расщепление: появились растения как с зелеными, так и морщинистыми семенами. Самое удивительное было то, что среди гибридов второго поколения оказались не только растения с желтыми гладкими и зелеными морщинистыми семенами. Также были желтые морщинистые и зеленые гладкие семена, т. е. произошла рекомбинация признаков, и получились такие комбинации, которые не встречались у исходных родительских форм.
Анализируя количественное соотношение разных семян F2, Мендель обнаружил следующее:
Если рассматривать каждый признак по отдельности, то он расщеплялся в отношении 3:1, как при моногибридном скрещивании. То есть на каждые три желтых семени приходилось одно зеленое, а на каждые 3 гладких — 1 морщинистое.
Появились растения с новыми комбинациями признаков.
Соотношение фенотипов было 9 : 3 : 3 : 1, где на девять желтых гладких семян гороха приходилось три желтых морщинистых, три зеленых гладких и одно зеленое морщинистое.
Третий закон Менделя хорошо иллюстрирует решетка Пеннета. Здесь в заголовках строк и столбцов пишутся возможные гаметы родителей (в данном случае гибридов первого поколения). Вероятность образования каждого типа гаметы составляет ¼. Также равновероятно различное их объединение в одну зиготу.
Мы видим, что образуется четыре фенотипа, два из которых ранее не существовали. Соотношение фенотипов 9 : 3 : 3 : 1. Количество разных генотипов и их соотношение более сложное:
AABB — 1 шт.
AABb — 2
AaBB — 2
AAbb — 1
AaBb — 4
Aabb — 2
aaBB — 1
aaBb — 2
aabb — 1
Получается 9 разных генотипов. Их соотношение: 4 : 2 : 2 : 2 : 2 : 1 : 1 : 1 : 1. При этом гетерозиготы встречаются чаще, а гомозиготы реже.
Если вернуться к тому, что каждый признак наследуется независимо, и по каждому наблюдается расщепление 3:1, то можно вычислить вероятность фенотипов по двум признакам разных аллелей, умножая вероятность проявления каждого аллеля (т. е. не обязательно пользоваться решеткой Пеннета). Так, вероятность гладких желтых семян будет равна ¾ × ¾ = 9/16, гладких зеленых – ¾ × ¼ = 3/16, морщинистых желтых – ¼ × ¾ = 3/16, морщинистых зеленых – ¼ × ¼ = 1/16. Таким образом, мы получаем то же соотношение фенотипов: 9:3:3:1.
Объясняется третий закон Менделя независимым расхождением гомологичных хромосом разных пар при первом делении мейоза. Хромосома, содержащая ген A, может с равной вероятностью уйти в одну клетку как с хромосомой, содержащей ген B, так и с хромосомой, содержащей ген b. Хромосома с геном A никак не привязана к хромосоме с геном B, хотя они обе и были унаследованы от одного родителя. Можно сказать, что в результате мейоза хромосомы перемешиваются. Количество различных их сочетаний вычисляется по формуле 2n, где n — это количество хромосом гаплоидного набора. Так, если у вида три пары хромосом, то количество различных их комбинаций будет равно 8 (23).
Когда не действует закон независимого наследования признаков
Третий закон Менделя, или закон независимого наследования признаков, действует только для генов, локализованных в разных хромосомах или расположенных в одной хромосоме, но достаточно далеко друг от друга.
В основном если гены находятся в одной хромосоме, то они наследуются совместно, т. е. проявляют сцепление между собой, и закон независимого наследования признаков уже не действует.
Например, если бы гены, отвечающие за окраску и форму семян гороха находились в одной хромосоме, то гибриды первого поколения могли бы образовывать гаметы только двух типов (AB и ab), так как в процессе мейоза независимо друг от друга расходятся родительские хромосомы, но не отдельные гены. В таком случае во втором поколении было бы расщепление 3:1 (три желтых гладких на одно зеленое морщинистое).
Однако не так все просто. Из-за существования в природе конъюгации (сближения) хромосом и кроссинговера (обмена участками хромосом) рекомбинируются и гены находящиеся в гомологичных хромосомах. Так, если хромосома с генами AB в процессе кроссинговера обменяется участком с геном B с гомологичной хромосомой, чей участок содержит ген b, то могут получиться новые гаметы (Ab и, например, aB). Процент таких рекомбинантных гамет будет меньше, чем если бы гены находились в разных хромосомах. При этом вероятность кроссинговера зависит от удаленности генов на хромосоме: чем дальше, тем вероятность больше.
«Гороховые» законы Грегора Менделя. Удивительная генетика
«Гороховые» законы Грегора Менделя

Основоположнику генетики Грегору Менделю выпал нелегкий жребий – он почти на полвека опередил свое время. А когда его вклад оценили наконец по достоинству и генетику открыли заново, далеко не все ученые были в восторге. Многие приняли новую науку откровенно в штыки и пренебрежительно именовали законы Менделя «гороховыми» законами.
Особенно не повезло генетике в Советской России. К началу 1930-х годов у нас сформировалась лучшая в мире генетическая школа. Но она была разрушена до основания, когда у руля отечественной биологии встали откровенные политические шарлатаны во главе с народным академиком Т. Д. Лысенко, а многие выдающиеся ученые, в том числе Н. И. Вавилов и С. С. Четвериков, бесследно сгинули в сталинских лагерях.
Мендель был современником Чарльза Дарвина. «Происхождение видов путем естественного отбора» увидело свет в 1859 году, а работа Менделя «Опыты над растительными гибридами» была опубликована в 1865 году.
Как известно, дарвинизм покоится на трех китах – наследственности, изменчивости и отборе, а одно из центральных положений фундаментального дарвиновского труда гласит, что наследственные вариации крайне незначительны, малозаметны, но со временем накапливаясь, они дают вполне ощутимые уклонения, которые, в конечном счете, и порождают виды. Сами по себе эти крохотные изменения (сегодня их называют мутациями, хотя в эпоху Дарвина такого термина не существовало) могут быть какими угодно по знаку – хоть вредными, хоть полезными, а сортирует их давление среды, выметая брак и бережно сохраняя ценные приобретения. В роли этого безличного селекционера выступает естественный отбор: особи с полезными наследственно обусловленными задатками активно развиваются, оставляя большое потомство, а организмы, обремененные вредными признаками, куда менее плодовиты и гибнут раньше.

Чарльз Дарвин
Как уже говорилось, «Происхождение видов путем естественного отбора» вышло из печати в 1859 году, а в 1867 году английский врач Флеминг Дженкин (1833–1885) выступил с критикой теории Дарвина, заявив, что малые вариации неизбежно будут находиться под угрозой исчезновения. И в самом деле: если организм приобретает новый признак (то есть некое отклонение в ту или иную сторону), а затем скрещивается с особью без всякого отклонения или с отклонением в сторону противоположную, то у потомков едва проклюнувшийся новый признак довольно быстро исчезнет или поменяет знак. Другими словами, любое наследственное уклонение будет неизбежно «разбавляться» в ряду поколений, пока от него почти ничего не останется.
Вот тут-то и появился монах-августинец Грегор Мендель, доказавший, что признак нельзя расколоть на произвольное количество фрагментов. Он неделим, подобно атому древних греков, и наследуется как единое целое, – доле?й или частиц признака в природе не существует. Более того, наследуются даже не сами признаки, а особые внутриклеточные структуры – гены, которые их кодируют, о чем, разумеется, в XIX веке не подозревали, но сумели сообразить, что за пестротой внешних проявлений должен прятаться некий вполне материальный носитель.
Грегор Мендель
В 1854–1868 годах Мендель преподавал физику и естественную историю в средней школе города Брюнне (ныне чешский город Брно), а на досуге выращивал горох в саду родного монастыря, причем руководствовался при этом соображениями высокой науки. Он был на редкость трудолюбив и аккуратен, и за восемь лет кропотливой работы вырастил около 10 тысяч гороховых кустов, прежде чем осмелился на лаконичную публикацию в местной прессе. Это историческое событие произошло в феврале 1865 года, и сегодня этот день по праву считается днем рождения современной генетики.
Менделю очень повезло с объектом. Его предшественники работали с материалом, который разнился между собой по нескольким признакам сразу, поэтому однозначно истолковать результат было весьма непросто. А вот горох имеет великое множество рас, которые отличаются друг от друга по одной-единственной паре альтернативных признаков. По сути дела, это чистые сорта, так что количественный анализ гибридов превращался в рутинную процедуру. Убедившись в однородности материала, Мендель проанализировал закономерность наследования как в тех случаях, когда родительские организмы отличались по одной альтернативной паре, так и в случаях, когда они различались по нескольким парам признаков. Скрещивание, в котором родительские особи анализируются по одной альтернативной паре признаков, называется моногибридным, по двум признакам – дигибридным, по многим – полигибридным.
Шарлотта Ауэрбах, профессор генетики Эдинбургского университета, пишет:
Мендель сознавал, что единственный путь к получению ясного ответа от природы – задать ей очень простой вопрос. Поэтому он взял для гибридизации сорта гороха, которые отличались друг от друга только по одному четко выраженному свойству или признаку, например, по форме семян или по окраске цветков. Это позволило ему четко классифицировать потомство в отношении только одного выбранного отличия; он мог так же точно подсчитывать, сколько растений попадает в один класс, а сколько в другой.
Грегор Мендель работал с горохом – самоопыляемым растением, поэтому у него была возможность проанализировать потомство каждой особи отдельно. Скрестив два сорта гороха – один с гладкими, а другой с морщинистыми семенами, – он обнаружил, что все потомство (так называемые гибриды первого поколения) имеет гладкие семена. Таким образом, у гибридов проявился лишь один из альтернативных родительских признаков, но и второй, как выяснилось впоследствии, не канул в небытие. Мендель назвал ведущий признак гибридов первого поколения доминантным, а тот, который временно спрятался, – рецессивным (от латинского recessus – отступление), а обнаруженную закономерность – правилом единообразия гибридов первого поколения.
После этого он отобрал гладкие гибридные семена, вырастил из них гороховые кусты, дал им созреть до цветения и самоопылиться. Получившиеся семена были уже вторым гибридным поколением, внуками исходных растений. Мендель тщательно их пересчитал и разбил на группы. Единообразие нарушилось: из 7324 семян 5474 оказались гладкими, а 1850 – морщинистыми. Будучи математиком, Мендель сразу же обратил внимание, что число 5474 почти втрое превышает число 1850. Расщепление признаков во втором поколении описывалось соотношением 3:1.
Аналогичные результаты обнаруживались и в других опытах, когда во внимание принимались иные признаки. Например, потомство растений с желтым и зеленым цветом семян всегда оказывалось желтого цвета, а при скрещивании между собой гибридов первого поколения непременно наблюдалось расщепление 3:1. Точно такая же картина наблюдалась при скрещивании растений с пурпурными и белыми цветками: сначала полное единообразие с доминированием пурпурной окраски, а затем известное нам расщепление в соотношении 3:1.
Мендель предположил, что в основе наследственности лежит некий материальный фактор, или носитель (о генах и хромосомах в ту пору не было даже речи). Попадая в зародышевые клетки, менделевские факторы комбинируются в процессе оплодотворения, а затем вновь расходятся, когда начинается очередной цикл гаметогенеза – производства половых клеток. Тем самым Мендель привнес в биологию точный расчет.
Но строгая теория великого чеха с математическими закономерностями настолько опережала тогдашние весьма расплывчатые представления о строении клетки и наследовании признаков, что оказалась совершенно невостребованной. Мысль о дис кретности наследственных факторов и свободном их комбинировании выглядела абсолютной ересью, поэтому его труд «Опыты над растительными гибридами», ставший впоследствии классическим, благополучно лег на полку.
Законы Менделя были «переоткрыты» спустя почти полвека, в 1900–1901 годах, совокупными усилиями трех ученых – голландца Гуго де Фриза, немца Карла Корренса и австрийца Эриха Чермака. И только тогда, с неприлично большим опозданием, научный мир по достоинству оценил безупречную логику эксперимента полузабытого чеха и красоту его расчетов.
Терминология Менделя заметно отличалась от современной. Сегодня его факторы, или задатки, называются генами. Это слово придумали в XX веке, когда поняли, что наследственные факторы расположены в хромосомах – особых структурах, лежащих внутри клеточного ядра. Разумеется, Мендель об этом понятия не имел, однако совершенно безукоризненно объяснил суть своего открытия. Чтобы получилось наблюдаемое распределение признаков, у родителей с самого начала должен быть двойной набор генов. А почему бы и нет? Тогда его результаты получают вполне естественное объяснение. Нужно только предположить, что у каждого из родителей имеется по два гена каждого сорта, а в зародыш попадает только по одному из них. И все тут же становится на свои места. Очевидно, что гены, отвечающие за окраску семян, неодинаковы и существуют в двух разновидностях – аллелях. Точно так же дело обстоит и с генами, управляющими формой семян.
Гуго де Фриз
Карл Корренс

Эрих Чермак
Чтобы не запутаться, обозначим эти гены, отвечающие за реализацию признака, буквами латинского алфавита. Гены, относящиеся к одной паре, принято обозначать одной и той же буквой, причем аллель доминантного состояния признака обозначают прописной буквой, а аллель рецессивного – строчной. Тогда аллель желтой окраски семян следует обозначить буквой А, аллель зеленой окраски – буквой а, гладкую аллель – буквой В, а морщинистую – буквой b. А теперь внимание: в клетке с двойным набором генов они могут быть как одинаковыми, так и разными – АА, аа или Аа. Организм с одинаковым аллельным вариантом называется гомозиготным, а с разным – гетерозиготным[1]. Итак, поскольку в соматической[2] клетке всегда содержатся оба варианта данного гена, особь, гомозиготную по доминантной аллели, мы запишем как АА, гомозиготную по рецессивной аллели, – как аа, а гетерозиготную – как Аа (если речь идет об окраске семян). Аналогично гомозигота по гладкой аллели запишется как ВВ, гомозигота по морщинистой аллели – как bb, а гетерозигота – как Bb. Важно отметить, что рецессивная аллель проявляется только в гомозиготном состоянии, а доминантная – как в гомозиготном, так и в гетерозиготном.
Как мы помним, каждый из родителей передает потомству только один вариант гена, поэтому у гомозиготного организма имеется только один тип половых клеток (гамет) – А или а. А вот у гетерозиготной особи половые клетки будут нести как доминантную аллель А, так и рецессивную а, причем их будет примерно поровну. Клетки с двойным набором генов (соматические) называются диплоидными, а половые клетки – гаплоидными, так как несут половинный набор. Забегая немного вперед, напомним значение терминов генотип и фенотип. Генотип – это совокупность генов (наследственных задатков), а фенотип – совокупность внешних признаков.
Таким образом, опыты по скрещиванию двух гомозиготных особей с разной окраской семян – желтой и зеленой – следует записать так: АА ? аа. Взаимодействующие между собой гаметы четырех типов – А, А, а, а – могут дать в итоге только один-единственный вариант – Аа, то есть потомство в первом поколении будет гетерозиготным по генотипу, а фенотипически – совершенно однородным (желтая окраска семян). Рецессивный ген а подавлен, скрыт, потому что его более сильный партнер А не дает ему проявиться.
Другими словами, гетерозигота в данном случае фенотипически неотличима от родительской гомозиготы с доминантным геном желтой окраски семян. Поэтому первое правило Менделя, или правило единообразия первого гибридного поколения, можно сформулировать так: при скрещивании гомозиготных особей, отличающихся друг от друга по одной паре альтернативных признаков, все потомство в первом поколении единообразно как по фенотипу, так и по генотипу.
А вот когда мы переходим к следующему этапу – скрещиваем гибриды первого поколения между собой, то получаем совершенно иную картину. Каждая гетерозигота (Аа) образует в равных количествах два типа гамет – половину с доминантными аллелями А и половину с рецессивными аллелями а. Яйцеклетка, несущая аллель А, может быть оплодотворена как сперматозоидом с аллелью А, так и сперматозоидом с аллелью а, причем вероятность этих событий примерно одинакова.
Точно так же дело обстоит и с яйцеклеткой, несущей аллель а: или а+А, или а+а. Поэтому на выходе мы получаем зиготы четырех типов – АА, Аа, Аа, аа. По фенотипу растения АА, Аа и Аа друг от друга неотличимы (единственное исключение – вариант аа), отсюда и возникает знаменитое менделевское расщепление 3:1, однако по генотипу расклад выглядит несколько иначе – 1АА: 2Аа: 1аа.
Итак, второй закон Менделя гласит: при скрещивании двух гетерозиготных особей, анализируемых по одной альтернативной паре признаков (то есть гибридов первого поколения), в потомстве ожидается расщепление по фенотипу в соотношении 3:1, а по генотипу – в соотношении 1:2:1.
Подытоживая сказанное, можно сделать следующие выводы:
1) аллельные гены, находящиеся в гетерозиготном состоянии, не изменяют друг друга;
2) при созревании гамет из гибридов образуется приблизительно равное число гамет с доминантными и рецессивными аллелями;
3) при оплодотворении мужские и женские гаметы, несущие доминантные и рецессивные аллели, свободно комбинируются.
Первый из этих выводов (несмешиваемость аллельных генов, которые находятся в гетерозиготном состоянии) Мендель назвал гипотезой чистоты гамет. Это явление, основанное на парности генов, расположенных в одних и тех же локусах[3] гомологичных хромосом, впоследствии получило исчерпывающее объяснение, когда цитологи как следует разобрались в строении клетки.
О дигибридном скрещивании (по двум парам признаков) мы здесь говорить не будем, а отметим только, что, например, форма семян наследуется независимо от их окраски, и расщепление признаков во втором поколении приобретает следующий вид: 9:3:3:1. Это третий и последний закон Менделя, или правило независимого комбинирования признаков.
Грегор Мендель был человеком, который пришел слишком рано, поэтому современники в большинстве своем отнеслись к его пионерским исследованиям как к забавному курьезу. Строгий количественный подход не впечатлил биологов XIX века. А вот если бы Дарвин внимательно прочитал замечательную работу чешского коллеги, теория происхождения видов раз и навсегда освободилась бы от невыносимого проклятия Дженкина – утраты вновь приобретенных признаков в ряду поколений.
Впрочем, существует и другое мнение: Дарвин был знаком с менделевской статьей, но просто-напросто не оценил ее по достоинству. Он полагал, что закономерности, открытые Менделем (правило единообразия гибридов первого поколения и расщепление в пропорции 3:1 во втором поколении), неприменимы к наследованию признаков в естественных популяциях. И надо сказать, что подобная точка зрения не лишена своей логики, потому что законы Менделя выполняются при скрещивании чистых линий, отличающихся только аллельным состоянием одного гена, а подобные примеры редко встречаются как в природе, так и в селекционной практике.
В теории происхождения видов было еще одно слабое место – вопрос о наследовании приобретенных признаков. Конечно, сегодня мы знаем, что приобретенные признаки не наследуются – об этом написано в школьном учебнике биологии. А вот Дарвин высказывался на эту тему весьма уклончиво, и тому были причины – в его время не было и не могло быть настоящей теории наследственности.
Август Вейсман
Точки над «i» расставил немецкий биолог Август Вейсман в начале ХХ века. На протяжении многих лет он рубил крысам хвосты и тщательно обмерял каждый новый приплод. Через его руки прошло двадцать два поколения несчастных грызунов, но хвосты у потомства упрямо продолжали вырастать до нормальной длины. Приобретенные признаки не наследовались.
Между прочим, этот хрестоматийный факт до сих пор плохо осознается людьми, далекими от биологии. Если спросить у человека с улицы, какой щенок скорее выучится трюкам на манеже – беспородный «дворянин» или благородный отпрыск из династии цирковых псов, – вам почти наверняка ответят, что, конечно же, цирковой – ведь все его предки работали на манеже. Разве могли не закрепиться в генах столь важные признаки?
Столь же безграмотными (с точки зрения биологии) выглядят рассуждения некоторых политиков и журналистов об особенностях национального характера на генетическом уровне (по поводу врожденной лености россиян, их горячей любви к твердой руке и проч.)
И почему-то никому не приходит в голову тот элементарный факт, что ребенок, увезенный за границу в младенческом возрасте, утрачивает даже малейшие следы российской ментальности…
После переоткрытия законов Менделя на рубеже XIX–XX веков генетика развивалась весьма бурно.
Было установлено, что гены находятся в хромосомах – небольших тельцах, лежащих попарно внутри клеточного ядра. Такие парные хромосомы называются гомологичными. Например, в соматических клетках человека содержится 46 хромосом, объединенных в 23 пары. Аллельные гены, кодирующие разные варианты одного и того же признака, расположены в одних и тех же локусах гомологичных хромосом. А вот половые клетки, образовавшиеся в результате особого варианта клеточного деления, содержат половинный набор хромосом. При слиянии половых клеток происходит восстановление исходного двойного набора, причем зародыш получает одну хромосому от отца, а другую от матери. Все в полном соответствии с законами Менделя.
В ХХ веке выкристаллизовалась хромосомная теория наследственности, а взаимодействие генов оказалось куда более сложным, чем это виделось Менделю. Было изучено взаимодействие аллельных (парных) генов и обнаружены такие явления, как неполное доминирование (когда ни один ген не является доминирующим, в результате в организме наблюдается влияние обоих генов) и сверхдоминирование (когда доминантная аллель ведет себя непоследовательно: в гетерозиготном состоянии проявляется ярко, а в гомозиготном – слабо).
При изучении неаллельных (непарных) генов было открыто много интересного. Если они влияют на один и тот же признак, усиливая его проявление, говорят о полимерии, а вот зависимость нескольких признаков от одного-единственного гена называется плейотропией. Например, довольно известная наследственная аномалия арахнодактилия («паучьи пальцы»), которая встречается при болезни Марфана и выражается в удлинении и утончении пальцев рук и ног, обусловлена геном, вызывающим, кроме того, патологию хрусталика в глазном яблоке.
Непарные гены умеют действовать и взаимодополняюще (комплементарно): каждый из них доминантен, но в отсутствие соседа молчит. Противоположное явление, когда непарная доминантная аллель подавляет действие другой доминантной аллели, называется эпистазом. Встречаются и множественные аллели, возникшие в результате многократного мутирования одного и того же хромосомного локуса.
Таким образом, помимо стандартной пары (доминантная плюс рецессивная версия гена) возникает третий член – промежуточная аллель, которая по отношению к доминантному гену ведет себя как рецессивная, а по отношению к рецессивному гену выступает в роли доминантной аллели. Между прочим, по такой схеме наследуются группы крови.
В XX столетии были открыты мутации (термин введен Гуго де Фризом) – внезапные скачкообразные изменения наследственных структур, поставляющие материал для естественного отбора. Различают мутации геномные (изменение числа хромосом), хромосомные (структурные изменения хромосом) и генные (их труднее всего распознать, поскольку они затрагивают отдельные гены).
Кроме того, мутации бывают соматическими и генеративными. Первые ограничиваются клетками тела, а вторые вмешиваются в генетический аппарат гамет и отражаются, таким образом, на последующих поколениях (если мутация происходит в клетках, из которых развиваются гаметы, или в самих половых клетках).
Наконец, существует фенотипическая (ненаследственная) изменчивость, которая не затрагивает генотип и называется модификационной. Она обусловлена средой обитания и является приспособительной. Даже на однояйцевых близнецов, которые имеют идентичный набор генов, условия среды могут повлиять весьма ощутимо. Если один из братьев занимается, предположим, гиревым спортом, а другой – легкой атлетикой, результат будет, что называется, налицо. Но их дети, разумеется, не унаследуют ни рельефной мускулатуры, ни феноменальной прыгучести, поскольку модификационные изменения по наследству не передаются.
а
б
Мутация ДНК: а – нормальная ДНК; б – измененная ДНК
Правда, у некоторых видов давление среды может приводить к возникновению длительных модификаций, которые закрепляются в ряду поколений. Так, например, при воздействии высокой или пониженной температуры на куколок колорадского жука окраска взрослых животных меняется. Этот признак удерживается в нескольких поколениях, а затем происходит возвращение прежней окраски.
По-видимому, в таких случаях работает цитоплазматическая наследственность: внешний фактор вызывает изменения в тех частях цитоплазмы (то есть гелеобразной внутриклеточной среды, в которой плавают органеллы – «внутренние органы» клетки; некоторые цитоплазматические структуры могут размножаться независимо от деления клетки), которые затем могут авторепродуцироваться.
Такой взгляд породил разного рода «катастрофические» сценарии, в которых масштабные изменения флоры и фауны связывают в первую очередь с природными катаклизмами (например, «астероидная» гипотеза вымирания динозавров). При этом совершенно упускается из виду тот общеизвестный факт, что эволюция вовсе не нуждается в катастрофах и может идти даже в условиях абсолютной стабильности, когда никаких изменений не происходит вовсе.
Выдающийся отечественный генетик С. С. Четвериков в статье «О некоторых моментах эволюционного процесса с точки зрения генетики», опубликованной еще в 1926 году, показал, что природные популяции несут в себе огромный запас ранее произошедших мутаций, буквально впитывают их, как губка впитывает воду, а значит, исходный материал у эволюции всегда в избытке.
И в самом деле: если большинство живых существ несут по два экземпляра каждого гена, а новые мутации, как правило, рецессивны (то есть внешне никак себя не проявляют), то под сходством внешнего облика представителей того или иного вида должно скрываться ошеломляющее генетическое разнообразие, накопившееся в длинном ряду поколений.
Данный текст является ознакомительным фрагментом.Читать книгу целиком
Поделитесь на страничкеСледующая глава >
МЕНДЕЛЯ ЗАКОНЫ • Большая российская энциклопедия
-

-
-
В книжной версии
Том 19. Москва, 2011, стр. 725-726
-
Скопировать библиографическую ссылку:



Авторы: В. С. Михеев
МЕ́НДЕЛЯ ЗАКО́НЫ, основные закономерности распределения наследственно детерминированных признаков в ряду последовательных поколений, установленные Г. Менделем. Эксперим. основой для формулировки М. з. послужили многолетние (1856–63) опыты по скрещиванию нескольких сортов гороха посевного (самоопыляющегося растения) с целью установления, как писал сам автор, «всеобщего закона образования и развития гибридов». Важную роль в разработке изучаемой Менделем проблемы сыграли: выбор объекта, преимущество которого для подобных опытов заключалось в многочисленности сортов (разновидностей) гороха, постоянстве их отличит. признаков, представленных качественными (альтернативными) формами, и одногодичности жизненного цикла; использование строгих методов подбора исходного материала, спец. схем скрещиваний; применение количественного учёта и математич. анализа полученных результатов. М. з. включают закон единообразия гибридов первого поколения, закон расщепления и закон независимого комбинирования признаков.

Рис. 1. Схема, иллюстрирующая на примере скрещивания двух сортов гороха (с жёлтыми и зелёными семенами) единообразие гибридов первого поколения (первый закон Менделя) и расщепление признаков у потомст…
Закон единообразия гибридов первого поколения (первый закон Менделя) утверждает, что у гибридов первого поколения от скрещивания форм, различающихся только по одному альтернативному признаку (напр., жёлтые или зелёные семена), проявляется признак только одного из родителей. Такое преобладание одного из признаков родителей у гибридов Мендель назвал доминированием, а соответствующий признак – доминантным (рис. 1). Альтернативные признаки, не проявляющиеся у гибридов первого поколения, он назвал рецессивными. Позднее было установлено, что полное доминирование одних признаков над другими представляет собой широко распространённое явление не только у растений, но и у животных, грибов, микрооганизмов. В некоторых случаях при скрещивании имеет место промежуточный фенотип (неполное доминирование). Так, у львиного зева цветки гибридных растений первого поколения от скрещивания родителей с малиновыми и белыми цветками всегда розовые. Бывает также, что в потомстве первого поколения проявляются признаки обоих родителей (т. н. кодоминирование). Напр., если один из родителей имеет группу крови А, а другой – В, то в крови их детей присутствуют антигены, характерные для обеих групп крови.
Закон расщепления (второй закон Менделя) гласит, что при скрещивании гибридов первого поколения между собой среди гибридов второго поколения в определённых соотношениях появляются особи с фенотипами исходных родительских форм и гибридов первого поколения. Так, в случае полного доминирования выявляются 75% особей с доминантным признаком и 25% с рецессивным, т. е. отношение числа особей с доминантным признаком к числу потомков с рецессивным признаком оказалось близким к 3 :1. Такое явление Мендель назвал расщеплением признаков. Впервые им был установлен факт, свидетельствующий о том, что растения с доминантными признаками, сходные по внешнему виду (по фенотипу), могут резко различаться по наследств. свойствам – по генотипу.

Рис. 2. Схема, иллюстрирующая независимое комбинирование признаков (третий закон Менделя). Наследование жёлтой (А) и зелёной (а) окраски семян, а также гладкой (В) и морщинистой (в) их формы. Аллели А…
Закон независимого комбинирования (наследования) признаков (третий закон Менделя) утверждает, что каждая пара альтернативных признаков (в совр. интерпретации – пара аллелей) наследуется в ряду поколений независимо друг от друга, в результате чего среди потомков второго поколения в определённом соотношении появляются особи с новыми (по отношению к родительским) комбинациями признаков (рис. 2). Так, при скрещивании исходных форм, различающихся по 2 признакам (напр., по окраске и форме семян), во втором поколении выявляются особи с 4 фенотипами в соотношении 9 : 3 : 3 : 1. При этом два фенотипа имеют «родительские» сочетания признаков, а остальные – новые. Этот закон основан на независимом поведении (расщеплении) нескольких пар гомологичных хромосом. Напр., при дигибридном скрещивании (участвуют особи, различающиеся по 2 парам признаков) это приводит к образованию у гибридов первого поколения 4 типов гамет (АВ, Ав, аВ, ав) и после образования зигот – к закономерному расщеплению по генотипу и соответственно по фенотипу.
Для выявления М. з. в их классич. форме необходимы: гомозиготность исходных форм, образование у гибридов гамет всех возможных типов в равных соотношениях, что обеспечивается правильным течением мейоза; одинаковая жизнеспособность гамет всех типов, равная вероятность встречи любых типов гамет при оплодотворении; одинаковая жизнеспособность зигот всех типов. Несоблюдение этих и некоторых других условий может приводить либо к отсутствию расщепления во втором поколении, либо к расщеплению в первом поколении, либо к искажению соотношения разл. гено- и фенотипов. М. з., вскрывшие дискретную, корпускулярную природу наследственности, имеют универсальный характер для всех диплоидных организмов, размножающихся половым способом. Для полиплоидов выявляют принципиально те же закономерности наследования, однако числовые соотношения гено- и фенотипич. классов отличаются от таковых у диплоидов. Соотношение классов изменяется и у диплоидов в случае сцепления генов («нарушение» третьего закона Менделя). В целом М. з. справедливы для аутосомных генов с полной пенетрантностью и постоянной экспрессивностью. При локализации генов в половых хромосомах или в ДНК органелл (пластиды, митохондрии) результаты реципрокных скрещиваний могут различаться и не следовать М. з., чего не наблюдается для генов, расположенных в аутосомах.
Современники Г. Менделя не смогли оценить важности сделанных им выводов, и лишь в 1900 М. з. были переоткрыты и правильно восприняты. Признание справедливости и значения М. з. в нач. 20 в. связано с определёнными успехами цитологии и формированием ядерной гипотезы наследственности. Механизмы, лежащие в основе М. з., были выяснены благодаря изучению путей образования половых клеток, в частности поведения хромосом в мейозе, и доказательству хромосомной теории наследственности. М. з. оказали огромное влияние на развитие классич. генетики. Они послужили основой для предположения о существовании в клетках (гаметах) наследств. факторов, контролирующих развитие признаков. Из М. з. следует, что эти факторы (гены) относительно постоянны, хотя и могут находиться в разл. состояниях, парны в соматич. клетках и единичны в гаметах, дискретны и могут вести себя независимо по отношению друг к другу.
Законы наследственности Г. Менделя
Биография

Грегор Иоганн Мендель (Gregor Johann Mendel) – выдающийся чешский естествоиспытатель. Он родился в Австрийской империи в простой крестьянской семье. При крещении он получил имя Иоганн.
Изучением природы мальчик увлекался с детства, когда еще работал, сперва помощником садовника, а затем – садовником. Проучившись некоторое время в институте Ольмюца, в философских классах, он в $1843$ году постригся в монахи и принял имя Грегор. Дальше с $1844$ по $1848$ год Грегор Мендель учился в Брюннском богословском институте и стал священником. Во время учебы он самостоятельно изучал многие науки, изучал в Венском университете естественную историю.
Именно в Вене Грегор Мендель увлекся исследованиями процессов гибридизации и статистическими соотношениями гибридов. Мендель уделял особое внимание вопросам изменений качественных признаков у растений. Объектом экспериментов он выбрал горох, который можно было вырастить в монастырском саду. Именно наблюдения за результатами этих исследований и легли в основу знаменитых «законов Менделя».
Воодушевленный первыми успехами, Мендель перенес свои эксперименты на растение семейства астровых (скрещивал разновидности ястребинки) и проводил скрещивания разновидностей пчел. Результаты экспериментов не совпали с результатами опытов с горохом. Тогда еще не знали, что механизм наследования признаков у этих растений и животных отличается от механизма наследования у гороха.
Замечание 1
Грегор Мендель был разочарован в биологической науке. После его назначения настоятелем монастыря, он больше не занимался наукой. Но его заслугой является то, что он впервые выявил и описал статистические закономерности наследования признаков у гибридов. Ознакомимся с ними детальнее.
Первый закон Менделя
Для облегчения учета результатов эксперимента Грегор Мендель избрал растения с четко отличающимися признаками. Это были цвет и форма семян.
Для начала он получил семена «чистых линий» растений. Эти семена при дальнейшем посеве и в результате самоопыления не давали расщепления признаков.
При скрещивании разных сортов гороха — с пурпурными цветками и с белыми цветками, в первом поколении гибридов Мендель получал все растения с пурпурными цветками. Аналогичными были результаты, когда ученый брал растения гороха с желтыми и зелеными семенами или семенами гладкой и морщинистой формы.
По результатам этих опытов Грегор Мендель вывел закон единообразия гибридов первого поколения, который мы знаем, как «первый закон Менделя». Сегодня он звучит так:
«При скрещивании двух гомозиготных организмов. которые относятся к чистым линиям и отличаются друг от друга по одной паре альтернативных проявлений определенного признака, всё первое поколение гибридов (F1) окажется полностью единообразным и будет нести проявление признака только одного из родителей».
Данный закон еще называют законом доминирования признаков. Он означает, что доминирующий признак появляется в фенотипе, подавляя рецессивный.
Второй закон Менделя
Проводя дальнейшие эксперименты с гибридами первого поколения, Мендель обнаружил, что при дальнейшем скрещивании гибридов первого поколения между собой гибриды второго поколений отличаются расщеплением признаков с устойчивым постоянством. Сегодня этот закон формулируют таким образом:
Определение 1
«После скрещивания двух гетерозиготных потомков первого поколения между собой, наблюдается расщепление во втором поколении в определенном числовом соотношении: по фенотипу $3:1$, по генотипу $1:2:1$».
Он получил название закона расщепления. Он означает, что рецессивный признак у гибридов первого поколения не исчезает, а лишь подавляется и потом проявляется во втором гибридном поколении.
Третий закон Менделя
В первых опытах Грегор Мендель принимал во внимание всего одну пару альтернативных признаков. Он заинтересовался вопросом, что если взять во внимание несколько признаков. Признаки начали комбинироваться между собой и поначалу вызвали у ученого замешательство. Но при более детальном рассмотрении, Менделю удалось вывести закономерность расщепления. Оказалось, что гибриды первого поколения однообразны, а во втором поколении признаки по фенотипу расщепляются в пропорции $9:3:3:1$, независимо от другого признака. Этот закон был назван законом независимого наследования. Сегодня его формулировка выглядит так:
Определение 2
«При скрещивании двух особей, которые отличаются друг от друга по нескольким парам (двум или более) альтернативных признаков, гены и соответствующие им признаки наследуются друг от друга независимо и могут комбинироваться во всех возможных сочетаниях (подобно как при моногибридном скрещивании)».

Закономерности, открытые Менделем предвосхитили начало новой науки – генетики.