Австралопитеки, австралопитековые. Их описание. Обезьяны, похожие на человека
Научный редактор АНТРОПОГЕНЕЗ.РУ, к.б.н., доцент кафедры антропологии биологического факультета МГУ им. Ломоносова Достающее звеноСпециально для портала «Антропогенез.РУ».
Авторский проект С.Дробышевского. Электронная книга даст читателям базовую информацию о том, что известно современной науке о древней родословной человека.
Древнейшими гоминидами обычно считают австралопитековых (Australopithecinae). Это была очень своеобразная группа, поскольку их одинаково точно можно описать и как двуногих обезьян, и как людей с обезьяньей головой. Сложность положения австралопитековых среди приматов заключается в том, что в их строении мозаично сочетаются признаки, характерные и для современных человекообразных обезьян, и для человека. Как относиться к такому сочетанию признаков?
Череп древнейшего австралопитека — Sahelanthropus tschadensis.
Череп получил прозвище «Тумай».
Источник: http://hominin.net/specimens/tm-266-01-060-1/
Самые ранние останки австралопитеков, найденные в Торос-Меналла (Республика Чад), датируются 6-7 млн. лет назад. Наиболее поздняя датировка была определена для находок массивных австралопитеков в Сварткрансе (Южная Африка) – 900 тыс. лет назад; это время существования уже намного более прогрессивных форм гоминид. Известны австралопитеки практически со всего указанного промежутка времени. Таким образом, период существования группы австралопитеков крайне велик.
Территория расселения австралопитеков также очень велика: вся Африка южнее Сахары и, возможно, некоторые территории севернее. Насколько известно, австралопитековые никогда не покидали пределов Африки. Находки вне этого континента, иногда приписывавшиеся австралопитекам (Тель-Убейдиа из Израиля, Мегантроп 1941 г. и Моджокерто с Явы), во всех случаях крайне фрагментарны и, следовательно, спорны.
AL 822-1 — череп самки австралопитека афарского (грацильный австралопитек).
Источник: William H. Kimbel and Yoel Rak. The cranial base of Australopithecus afarensis: new insights from the female skull.
Phil. Trans. R. Soc. B 2010 365, 3365-3376
Австралопитеков можно разделить на три основных группы, сравнительно последовательно сменявшие друг друга во времени, в каждой из них выделяется несколько видов:
Ранние австралопитеки – существовали с 7 до 4 млн. лет назад, обладали наиболее примитивным строением. Выделяют несколько родов и видов ранних австралопитеков.
лет назад, обладали наиболее примитивным строением. Выделяют несколько родов и видов ранних австралопитеков.
Грацильные австралопитеки – существовали с 4 до 2,5 млн. лет назад, имели сравнительно небольшие размеры и умеренные пропорции. Обычно выделяют один род Australopithecus c несколькими видами.
Массивные австралопитеки – существовали с 2,5 до 1 млн. лет назад, были очень массивно сложенными специализированными формами с крайне развитыми челюстями, маленькими передними и огромными задними зубами. Массивные австралопитеки выделяются в самостоятельный род Paranthropus
с тремя видами.Относительно их детальной таксономии существует множество точек зрения; можно считать твёрдо установленным факт видовых различий по крайней мере между грацильными и массивными австралопитеками. Таксономические взаимоотношения внутри этих групп, даже между синхронными группами Восточной и Южной Африки – неясны.
Одновременное сосуществование разных «хороших» видов австралопитеков на одной территории не было твёрдо доказано ни для одного местонахождения, хотя предположений на этот счет выдвигалось много. Однако сосуществование австралопитеков с представителями «эугоминид» (или «ранними Homo«) не вызывает сомнения, по крайней мере для Восточной Африки.
Однако сосуществование австралопитеков с представителями «эугоминид» (или «ранними Homo«) не вызывает сомнения, по крайней мере для Восточной Африки.
Австралопитеки — это… Что такое Австралопитеки?
Австралопите́ки (от лат. australis — южный и др.-греч. πίθηκος — обезьяна) — группа ископаемых высших приматов, кости которых впервые были обнаружены в пустыне Калахари (Южная Африка) в 1924 году, а затем в Восточной и Центральной Африке. Австралопитеками принято считать всех двуногоходящих обезьян, с объёмом мозга до 880 см³. Вероятно являются предками рода Люди
Происхождение, биология и поведение
Близкие к австралопитекам приматы были распространены в Передней, Южной и Юго-Восточной Азии. Австралопитеки жили в плиоцене примерно с 4 млн лет назад, до менее миллиона лет назад. На временной шкале хорошо прослеживаются 3 длинных эпохи основных видов, примерно по миллиону лет на вид. Большинство видов австралопитеков были всеядны, однако были подвиды специализировавшиеся на растительной пище. Предком основного вида скорее всего был вид anamensis, а первым основным видом известным на данный момент стал вид afarensis, который просуществовал примерно 1 млн лет. По-видимому, эти существа были не более чем обезьянами, по-человечески перемещавшимися на двух ногах, хотя и сгорбленно. Возможно, под конец они умели пользоваться подручными камнями для раскалывания, к примеру, орехов. Считается, что afarensis в конце разделился на два подвида: первая ветвь пошла к очеловечиванию и homo habilis, вторая — продолжила совершенствоваться в австралопитеках, образовав новый вид africanus. У africanus были немного менее развиты конечности, чем у afarensis, но зато они научились пользоваться подручными камнями, палками, и острыми осколками костей, и, в свою очередь, ещё через миллион лет образовали два новых высших и последних известных подвида австралопитеков boisei и robustus, которые просуществовали вплоть до 900 тыс. лет до н. э. и уже могли самостоятельно изготавливать простейшие костяные и деревянные орудия.
Предком основного вида скорее всего был вид anamensis, а первым основным видом известным на данный момент стал вид afarensis, который просуществовал примерно 1 млн лет. По-видимому, эти существа были не более чем обезьянами, по-человечески перемещавшимися на двух ногах, хотя и сгорбленно. Возможно, под конец они умели пользоваться подручными камнями для раскалывания, к примеру, орехов. Считается, что afarensis в конце разделился на два подвида: первая ветвь пошла к очеловечиванию и homo habilis, вторая — продолжила совершенствоваться в австралопитеках, образовав новый вид africanus. У africanus были немного менее развиты конечности, чем у afarensis, но зато они научились пользоваться подручными камнями, палками, и острыми осколками костей, и, в свою очередь, ещё через миллион лет образовали два новых высших и последних известных подвида австралопитеков boisei и robustus, которые просуществовали вплоть до 900 тыс. лет до н. э. и уже могли самостоятельно изготавливать простейшие костяные и деревянные орудия.
С точки зрения таксономии, австралопитеков относят к семейству гоминид (включающему также разные виды людей и современных крупных человекообразных обезьян). Вопрос о том, были ли какие-либо австралопитеки предками людей, или они представляют собой «сестринскую» по отношению к людям группу, не выяснен до конца.
Анатомия
Череп самки Australopithecus africanus | ||||||||||
С человеком австралопитеков сближает слабое развитие челюстей, отсутствие крупных выступающих клыков, хватательная кисть с развитым большим пальцем, опорная стопа и строение таза, приспособленное для прямохождения.

Развитие форм внутри рода
Основным кандидатом на место предка австралопитеков является род Ardipithecus. При этом самый древний из представителей нового рода, Australopithecus anamensis, произошёл непосредственно от Ardipithecus ramidus 4,4—4,1 миллиона лет назад, а 3,6 миллиона лет назад дал начало Australopithecus afarensis, к которому принадлежит знаменитая Люси. С находкой в 1985 году так называемого «чёрного черепа», который был очень похож на Paranthropus boisei, с характерным костяным гребнем, но при этом был на 2,5 млн старше, в родословной австралопитеков появилась официальная неопределённость, поскольку хоть и результаты анализов могут сильно меняться в зависимости от множества обстоятельств и среды, где находился череп, и, как водится, будут ещё десятилетиями перепроверяться десятки раз, но на текущий момент получается, что Paranthropus boisei не мог произойти от Australopithecus africanus, поскольку жил до них, и как минимум жил в одно время с Australopithecus afarensis, и, соответственно, тоже не мог от них произойти, если, конечно, не учитывать гипотезу, что парантропные формы австралопитеков и австалопитеки являются самцами и самками одного и того же вида.
Известные формы
Ранее к роду Australopithecus причисляли ещё трёх представителей, однако в настоящее время их принято выделять в особый род парантропов (Paranthropus).
Место в эволюции гоминид
Реконструкция самки Australopithecus afarensis | ||||||||||
Род Australopithecus считается предком по меньшей мере двух групп гоминид: парантропов и людей. Хотя по уровню интеллекта австралопитеки мало отличались от обезьян, они были прямоходящими, в то время как большинство обезьян являются четвероногими. Таким образом, прямохождение предшествовало развитию интеллекта у людей, а не наоборот, как ранее предполагалось.
Как австралопитеки перешли к прямохождению, пока не ясно. Среди рассматриваемых причин называют необходимость хватать объекты, такие как пищу и детенышей, передними лапами, и осматривать окрестности поверх высокой травы в поисках пищи или чтобы вовремя заметить опасность. Высказывается также предположение, что общие предки прямоходящих гоминид (в том числе людей и австралопитеков) жили на мелководье, и питались мелкими водными обитателями, а прямохождение сформировалось как адаптация к передвижению по мелководью. В пользу этой версии говорит ряд анатомо-физиологических и этологических особенностей, в частности — способность людей произвольно задерживать дыхание, на что способны не все плавающие животные.
Высказывается также предположение, что общие предки прямоходящих гоминид (в том числе людей и австралопитеков) жили на мелководье, и питались мелкими водными обитателями, а прямохождение сформировалось как адаптация к передвижению по мелководью. В пользу этой версии говорит ряд анатомо-физиологических и этологических особенностей, в частности — способность людей произвольно задерживать дыхание, на что способны не все плавающие животные.
Согласно данным генетики, признаки прямохождения появились у некоторых вымерших видов обезьян ещё около 6 млн лет назад, в эпоху расхождения между людьми и шимпанзе. Это значит, что не только сами австралопитеки, но и вид, который был их предком, например, ардипитек, уже мог быть прямоходящим. Возможно, прямохождение было элементом адаптации к жизни на деревьях. Современные орангутаны используют все четыре лапы для перемещения только по толстым сучьям, в то время как к более тонким ветвям либо цепляются снизу, либо идут по ним на задних лапах, готовясь передними уцепиться за другие ветви повыше или балансируя для устойчивости. Такая тактика позволяет им приблизиться к плодам, расположенным далеко от ствола, или прыгать с одного дерева на другое. Изменения климата, произошедшие 11-12 млн лет назад, привели к сокращению лесных массивов в Африке и появлению больших открытых пространств, что и могло подтолкнуть предков австралопитеков к переходу к прямохождению по земле. В отличие от них предки современных шимпанзе и горилл специализировались на лазании по вертикальным стволам и лианам, чем обусловлена их кривоногая и косолапая походка на земле. Тем не менее, люди унаследовали много общего с этими обезьянами, включая строение костей рук, усиленных для хождения с опорой на костяшки пальцев[3][4].
Такая тактика позволяет им приблизиться к плодам, расположенным далеко от ствола, или прыгать с одного дерева на другое. Изменения климата, произошедшие 11-12 млн лет назад, привели к сокращению лесных массивов в Африке и появлению больших открытых пространств, что и могло подтолкнуть предков австралопитеков к переходу к прямохождению по земле. В отличие от них предки современных шимпанзе и горилл специализировались на лазании по вертикальным стволам и лианам, чем обусловлена их кривоногая и косолапая походка на земле. Тем не менее, люди унаследовали много общего с этими обезьянами, включая строение костей рук, усиленных для хождения с опорой на костяшки пальцев[3][4].
Не исключено также, что австралопитеки не были прямыми предками людей, а представляли собой тупиковую ветвь эволюции. К таким выводам подталкивают, в частности, недавние находки сахелантропа, ещё более древней человекообразной обезьяны, которая была больше похожа на Homo erectus, чем австралопитеки. В 2008 году был открыт новый вид австралопитеков, A. sediba, живший в Африке менее двух миллионов лет назад. Хотя по отдельным морфологическим признакам он ближе к людям, чем более древние виды австралопитеков, что дало основание его открывателям объявить его переходной формой от австралопитеков к людям, одновременно с ним, по-видимому, уже существовали первые представители рода Homo[5], такие, как человек рудольфский, что исключает возможность того, что этот вид австралопитеков мог быть предком современного человека.
В 2008 году был открыт новый вид австралопитеков, A. sediba, живший в Африке менее двух миллионов лет назад. Хотя по отдельным морфологическим признакам он ближе к людям, чем более древние виды австралопитеков, что дало основание его открывателям объявить его переходной формой от австралопитеков к людям, одновременно с ним, по-видимому, уже существовали первые представители рода Homo[5], такие, как человек рудольфский, что исключает возможность того, что этот вид австралопитеков мог быть предком современного человека.
Большинство видов австралопитеков использовали орудия труда не более современных обезьян. Известно, что шимпанзе и гориллы способны колоть орехи камнями, использовать палочки для извлечения термитов и дубинки для охоты. Насколько часто австралопитеки охотились — вопрос спорный, так как их ископаемые останки редко ассоциируют с останками убитых животных.
См. также
Примечания
Ссылки
Австралопитеки — это.
 .. Что такое Австралопитеки?
.. Что такое Австралопитеки?Австралопите́ки (от лат. australis — южный и др.-греч. πίθηκος — обезьяна) — группа ископаемых высших приматов, кости которых впервые были обнаружены в пустыне Калахари (Южная Африка) в 1924 году, а затем в Восточной и Центральной Африке. Австралопитеками принято считать всех двуногоходящих обезьян, с объёмом мозга до 880 см³. Вероятно являются предками рода Люди[1].
Происхождение, биология и поведение
Близкие к австралопитекам приматы были распространены в Передней, Южной и Юго-Восточной Азии. Австралопитеки жили в плиоцене примерно с 4 млн лет назад, до менее миллиона лет назад. На временной шкале хорошо прослеживаются 3 длинных эпохи основных видов, примерно по миллиону лет на вид. Большинство видов австралопитеков были всеядны, однако были подвиды специализировавшиеся на растительной пище. Предком основного вида скорее всего был вид anamensis, а первым основным видом известным на данный момент стал вид afarensis, который просуществовал примерно 1 млн лет. По-видимому, эти существа были не более чем обезьянами, по-человечески перемещавшимися на двух ногах, хотя и сгорбленно. Возможно, под конец они умели пользоваться подручными камнями для раскалывания, к примеру, орехов. Считается, что afarensis в конце разделился на два подвида: первая ветвь пошла к очеловечиванию и homo habilis, вторая — продолжила совершенствоваться в австралопитеках, образовав новый вид africanus. У africanus были немного менее развиты конечности, чем у afarensis, но зато они научились пользоваться подручными камнями, палками, и острыми осколками костей, и, в свою очередь, ещё через миллион лет образовали два новых высших и последних известных подвида австралопитеков boisei и robustus, которые просуществовали вплоть до 900 тыс. лет до н. э. и уже могли самостоятельно изготавливать простейшие костяные и деревянные орудия. Несмотря на это, большинство австралопитеков входило в пищевую цепочку более прогрессивных людей обогнавших их в развитии по другим веткам эволюции, и с которыми они пересекались по времени, хотя продолжительность совместного существования указывает, что были и периоды мирного совместного существования.
По-видимому, эти существа были не более чем обезьянами, по-человечески перемещавшимися на двух ногах, хотя и сгорбленно. Возможно, под конец они умели пользоваться подручными камнями для раскалывания, к примеру, орехов. Считается, что afarensis в конце разделился на два подвида: первая ветвь пошла к очеловечиванию и homo habilis, вторая — продолжила совершенствоваться в австралопитеках, образовав новый вид africanus. У africanus были немного менее развиты конечности, чем у afarensis, но зато они научились пользоваться подручными камнями, палками, и острыми осколками костей, и, в свою очередь, ещё через миллион лет образовали два новых высших и последних известных подвида австралопитеков boisei и robustus, которые просуществовали вплоть до 900 тыс. лет до н. э. и уже могли самостоятельно изготавливать простейшие костяные и деревянные орудия. Несмотря на это, большинство австралопитеков входило в пищевую цепочку более прогрессивных людей обогнавших их в развитии по другим веткам эволюции, и с которыми они пересекались по времени, хотя продолжительность совместного существования указывает, что были и периоды мирного совместного существования.
С точки зрения таксономии, австралопитеков относят к семейству гоминид (включающему также разные виды людей и современных крупных человекообразных обезьян). Вопрос о том, были ли какие-либо австралопитеки предками людей, или они представляют собой «сестринскую» по отношению к людям группу, не выяснен до конца.
Анатомия
Череп самки Australopithecus africanus | ||||||||||
С человеком австралопитеков сближает слабое развитие челюстей, отсутствие крупных выступающих клыков, хватательная кисть с развитым большим пальцем, опорная стопа и строение таза, приспособленное для прямохождения. Головной мозг относительно крупный (530 см³), но по строению мало отличающийся от мозга современных человекообразных обезьян. По объёму он составлял не более 35 % от средних размеров мозга современного человека. Размеры тела также были невелики, не более 120—140 см в высоту, телосложение стройное. Предполагается, что самцы были существенно крупнее самок, чем у современных гоминид. Например, у современных людей мужчины в среднем лишь на 15 % крупнее женщин, в то время как у австралопитеков они могли быть на 50 % выше и тяжелее[2], что порождает дискуссии о принципиальной возможности столь сильного полового диморфизма у этого рода гоминид. Одним из основных характерных признаков для парантропов является костяной стреловидный гребень на черепе, присущий самцам современных горилл, поэтому нельзя до конца исключить, что робустерные/парантропные формы австралопитеков являются самцами, а грацильные — самками, альтернативным объяснением может быть отнесение форм разных размеров к разным видам или подвидам.
Размеры тела также были невелики, не более 120—140 см в высоту, телосложение стройное. Предполагается, что самцы были существенно крупнее самок, чем у современных гоминид. Например, у современных людей мужчины в среднем лишь на 15 % крупнее женщин, в то время как у австралопитеков они могли быть на 50 % выше и тяжелее[2], что порождает дискуссии о принципиальной возможности столь сильного полового диморфизма у этого рода гоминид. Одним из основных характерных признаков для парантропов является костяной стреловидный гребень на черепе, присущий самцам современных горилл, поэтому нельзя до конца исключить, что робустерные/парантропные формы австралопитеков являются самцами, а грацильные — самками, альтернативным объяснением может быть отнесение форм разных размеров к разным видам или подвидам.
Развитие форм внутри рода
Основным кандидатом на место предка австралопитеков является род Ardipithecus. При этом самый древний из представителей нового рода, Australopithecus anamensis, произошёл непосредственно от Ardipithecus ramidus 4,4—4,1 миллиона лет назад, а 3,6 миллиона лет назад дал начало Australopithecus afarensis, к которому принадлежит знаменитая Люси. С находкой в 1985 году так называемого «чёрного черепа», который был очень похож на Paranthropus boisei, с характерным костяным гребнем, но при этом был на 2,5 млн старше, в родословной австралопитеков появилась официальная неопределённость, поскольку хоть и результаты анализов могут сильно меняться в зависимости от множества обстоятельств и среды, где находился череп, и, как водится, будут ещё десятилетиями перепроверяться десятки раз, но на текущий момент получается, что Paranthropus boisei не мог произойти от Australopithecus africanus, поскольку жил до них, и как минимум жил в одно время с Australopithecus afarensis, и, соответственно, тоже не мог от них произойти, если, конечно, не учитывать гипотезу, что парантропные формы австралопитеков и австалопитеки являются самцами и самками одного и того же вида.
С находкой в 1985 году так называемого «чёрного черепа», который был очень похож на Paranthropus boisei, с характерным костяным гребнем, но при этом был на 2,5 млн старше, в родословной австралопитеков появилась официальная неопределённость, поскольку хоть и результаты анализов могут сильно меняться в зависимости от множества обстоятельств и среды, где находился череп, и, как водится, будут ещё десятилетиями перепроверяться десятки раз, но на текущий момент получается, что Paranthropus boisei не мог произойти от Australopithecus africanus, поскольку жил до них, и как минимум жил в одно время с Australopithecus afarensis, и, соответственно, тоже не мог от них произойти, если, конечно, не учитывать гипотезу, что парантропные формы австралопитеков и австалопитеки являются самцами и самками одного и того же вида.
Известные формы
Ранее к роду Australopithecus причисляли ещё трёх представителей, однако в настоящее время их принято выделять в особый род парантропов (Paranthropus).
Место в эволюции гоминид
Реконструкция самки Australopithecus afarensis | ||||||||||
Род Australopithecus считается предком по меньшей мере двух групп гоминид: парантропов и людей. Хотя по уровню интеллекта австралопитеки мало отличались от обезьян, они были прямоходящими, в то время как большинство обезьян являются четвероногими. Таким образом, прямохождение предшествовало развитию интеллекта у людей, а не наоборот, как ранее предполагалось.
Как австралопитеки перешли к прямохождению, пока не ясно. Среди рассматриваемых причин называют необходимость хватать объекты, такие как пищу и детенышей, передними лапами, и осматривать окрестности поверх высокой травы в поисках пищи или чтобы вовремя заметить опасность. Высказывается также предположение, что общие предки прямоходящих гоминид (в том числе людей и австралопитеков) жили на мелководье, и питались мелкими водными обитателями, а прямохождение сформировалось как адаптация к передвижению по мелководью. В пользу этой версии говорит ряд анатомо-физиологических и этологических особенностей, в частности — способность людей произвольно задерживать дыхание, на что способны не все плавающие животные.
В пользу этой версии говорит ряд анатомо-физиологических и этологических особенностей, в частности — способность людей произвольно задерживать дыхание, на что способны не все плавающие животные.
Согласно данным генетики, признаки прямохождения появились у некоторых вымерших видов обезьян ещё около 6 млн лет назад, в эпоху расхождения между людьми и шимпанзе. Это значит, что не только сами австралопитеки, но и вид, который был их предком, например, ардипитек, уже мог быть прямоходящим. Возможно, прямохождение было элементом адаптации к жизни на деревьях. Современные орангутаны используют все четыре лапы для перемещения только по толстым сучьям, в то время как к более тонким ветвям либо цепляются снизу, либо идут по ним на задних лапах, готовясь передними уцепиться за другие ветви повыше или балансируя для устойчивости. Такая тактика позволяет им приблизиться к плодам, расположенным далеко от ствола, или прыгать с одного дерева на другое. Изменения климата, произошедшие 11-12 млн лет назад, привели к сокращению лесных массивов в Африке и появлению больших открытых пространств, что и могло подтолкнуть предков австралопитеков к переходу к прямохождению по земле. В отличие от них предки современных шимпанзе и горилл специализировались на лазании по вертикальным стволам и лианам, чем обусловлена их кривоногая и косолапая походка на земле. Тем не менее, люди унаследовали много общего с этими обезьянами, включая строение костей рук, усиленных для хождения с опорой на костяшки пальцев[3][4].
В отличие от них предки современных шимпанзе и горилл специализировались на лазании по вертикальным стволам и лианам, чем обусловлена их кривоногая и косолапая походка на земле. Тем не менее, люди унаследовали много общего с этими обезьянами, включая строение костей рук, усиленных для хождения с опорой на костяшки пальцев[3][4].
Не исключено также, что австралопитеки не были прямыми предками людей, а представляли собой тупиковую ветвь эволюции. К таким выводам подталкивают, в частности, недавние находки сахелантропа, ещё более древней человекообразной обезьяны, которая была больше похожа на Homo erectus, чем австралопитеки. В 2008 году был открыт новый вид австралопитеков, A. sediba, живший в Африке менее двух миллионов лет назад. Хотя по отдельным морфологическим признакам он ближе к людям, чем более древние виды австралопитеков, что дало основание его открывателям объявить его переходной формой от австралопитеков к людям, одновременно с ним, по-видимому, уже существовали первые представители рода Homo[5], такие, как человек рудольфский, что исключает возможность того, что этот вид австралопитеков мог быть предком современного человека.
Большинство видов австралопитеков использовали орудия труда не более современных обезьян. Известно, что шимпанзе и гориллы способны колоть орехи камнями, использовать палочки для извлечения термитов и дубинки для охоты. Насколько часто австралопитеки охотились — вопрос спорный, так как их ископаемые останки редко ассоциируют с останками убитых животных.
См. также
Примечания
Ссылки
Австралопитеки — это… Что такое Австралопитеки?
Австралопите́ки (от лат. australis — южный и др.-греч. πίθηκος — обезьяна) — группа ископаемых высших приматов, кости которых впервые были обнаружены в пустыне Калахари (Южная Африка) в 1924 году, а затем в Восточной и Центральной Африке. Австралопитеками принято считать всех двуногоходящих обезьян, с объёмом мозга до 880 см³. Вероятно являются предками рода Люди[1].
Происхождение, биология и поведение
Близкие к австралопитекам приматы были распространены в Передней, Южной и Юго-Восточной Азии. Австралопитеки жили в плиоцене примерно с 4 млн лет назад, до менее миллиона лет назад. На временной шкале хорошо прослеживаются 3 длинных эпохи основных видов, примерно по миллиону лет на вид. Большинство видов австралопитеков были всеядны, однако были подвиды специализировавшиеся на растительной пище. Предком основного вида скорее всего был вид anamensis, а первым основным видом известным на данный момент стал вид afarensis, который просуществовал примерно 1 млн лет. По-видимому, эти существа были не более чем обезьянами, по-человечески перемещавшимися на двух ногах, хотя и сгорбленно. Возможно, под конец они умели пользоваться подручными камнями для раскалывания, к примеру, орехов. Считается, что afarensis в конце разделился на два подвида: первая ветвь пошла к очеловечиванию и homo habilis, вторая — продолжила совершенствоваться в австралопитеках, образовав новый вид africanus. У africanus были немного менее развиты конечности, чем у afarensis, но зато они научились пользоваться подручными камнями, палками, и острыми осколками костей, и, в свою очередь, ещё через миллион лет образовали два новых высших и последних известных подвида австралопитеков boisei и robustus, которые просуществовали вплоть до 900 тыс.
Австралопитеки жили в плиоцене примерно с 4 млн лет назад, до менее миллиона лет назад. На временной шкале хорошо прослеживаются 3 длинных эпохи основных видов, примерно по миллиону лет на вид. Большинство видов австралопитеков были всеядны, однако были подвиды специализировавшиеся на растительной пище. Предком основного вида скорее всего был вид anamensis, а первым основным видом известным на данный момент стал вид afarensis, который просуществовал примерно 1 млн лет. По-видимому, эти существа были не более чем обезьянами, по-человечески перемещавшимися на двух ногах, хотя и сгорбленно. Возможно, под конец они умели пользоваться подручными камнями для раскалывания, к примеру, орехов. Считается, что afarensis в конце разделился на два подвида: первая ветвь пошла к очеловечиванию и homo habilis, вторая — продолжила совершенствоваться в австралопитеках, образовав новый вид africanus. У africanus были немного менее развиты конечности, чем у afarensis, но зато они научились пользоваться подручными камнями, палками, и острыми осколками костей, и, в свою очередь, ещё через миллион лет образовали два новых высших и последних известных подвида австралопитеков boisei и robustus, которые просуществовали вплоть до 900 тыс. лет до н. э. и уже могли самостоятельно изготавливать простейшие костяные и деревянные орудия. Несмотря на это, большинство австралопитеков входило в пищевую цепочку более прогрессивных людей обогнавших их в развитии по другим веткам эволюции, и с которыми они пересекались по времени, хотя продолжительность совместного существования указывает, что были и периоды мирного совместного существования.
лет до н. э. и уже могли самостоятельно изготавливать простейшие костяные и деревянные орудия. Несмотря на это, большинство австралопитеков входило в пищевую цепочку более прогрессивных людей обогнавших их в развитии по другим веткам эволюции, и с которыми они пересекались по времени, хотя продолжительность совместного существования указывает, что были и периоды мирного совместного существования.
С точки зрения таксономии, австралопитеков относят к семейству гоминид (включающему также разные виды людей и современных крупных человекообразных обезьян). Вопрос о том, были ли какие-либо австралопитеки предками людей, или они представляют собой «сестринскую» по отношению к людям группу, не выяснен до конца.
Анатомия
Череп самки Australopithecus africanus | ||||||||||
С человеком австралопитеков сближает слабое развитие челюстей, отсутствие крупных выступающих клыков, хватательная кисть с развитым большим пальцем, опорная стопа и строение таза, приспособленное для прямохождения. Головной мозг относительно крупный (530 см³), но по строению мало отличающийся от мозга современных человекообразных обезьян. По объёму он составлял не более 35 % от средних размеров мозга современного человека. Размеры тела также были невелики, не более 120—140 см в высоту, телосложение стройное. Предполагается, что самцы были существенно крупнее самок, чем у современных гоминид. Например, у современных людей мужчины в среднем лишь на 15 % крупнее женщин, в то время как у австралопитеков они могли быть на 50 % выше и тяжелее[2], что порождает дискуссии о принципиальной возможности столь сильного полового диморфизма у этого рода гоминид. Одним из основных характерных признаков для парантропов является костяной стреловидный гребень на черепе, присущий самцам современных горилл, поэтому нельзя до конца исключить, что робустерные/парантропные формы австралопитеков являются самцами, а грацильные — самками, альтернативным объяснением может быть отнесение форм разных размеров к разным видам или подвидам.
Головной мозг относительно крупный (530 см³), но по строению мало отличающийся от мозга современных человекообразных обезьян. По объёму он составлял не более 35 % от средних размеров мозга современного человека. Размеры тела также были невелики, не более 120—140 см в высоту, телосложение стройное. Предполагается, что самцы были существенно крупнее самок, чем у современных гоминид. Например, у современных людей мужчины в среднем лишь на 15 % крупнее женщин, в то время как у австралопитеков они могли быть на 50 % выше и тяжелее[2], что порождает дискуссии о принципиальной возможности столь сильного полового диморфизма у этого рода гоминид. Одним из основных характерных признаков для парантропов является костяной стреловидный гребень на черепе, присущий самцам современных горилл, поэтому нельзя до конца исключить, что робустерные/парантропные формы австралопитеков являются самцами, а грацильные — самками, альтернативным объяснением может быть отнесение форм разных размеров к разным видам или подвидам.
Развитие форм внутри рода
Основным кандидатом на место предка австралопитеков является род Ardipithecus. При этом самый древний из представителей нового рода, Australopithecus anamensis, произошёл непосредственно от Ardipithecus ramidus 4,4—4,1 миллиона лет назад, а 3,6 миллиона лет назад дал начало Australopithecus afarensis, к которому принадлежит знаменитая Люси. С находкой в 1985 году так называемого «чёрного черепа», который был очень похож на Paranthropus boisei, с характерным костяным гребнем, но при этом был на 2,5 млн старше, в родословной австралопитеков появилась официальная неопределённость, поскольку хоть и результаты анализов могут сильно меняться в зависимости от множества обстоятельств и среды, где находился череп, и, как водится, будут ещё десятилетиями перепроверяться десятки раз, но на текущий момент получается, что Paranthropus boisei не мог произойти от Australopithecus africanus, поскольку жил до них, и как минимум жил в одно время с Australopithecus afarensis, и, соответственно, тоже не мог от них произойти, если, конечно, не учитывать гипотезу, что парантропные формы австралопитеков и австалопитеки являются самцами и самками одного и того же вида.
Известные формы
Ранее к роду Australopithecus причисляли ещё трёх представителей, однако в настоящее время их принято выделять в особый род парантропов (Paranthropus).
Место в эволюции гоминид
Реконструкция самки Australopithecus afarensis | ||||||||||
Род Australopithecus считается предком по меньшей мере двух групп гоминид: парантропов и людей. Хотя по уровню интеллекта австралопитеки мало отличались от обезьян, они были прямоходящими, в то время как большинство обезьян являются четвероногими. Таким образом, прямохождение предшествовало развитию интеллекта у людей, а не наоборот, как ранее предполагалось.
Как австралопитеки перешли к прямохождению, пока не ясно. Среди рассматриваемых причин называют необходимость хватать объекты, такие как пищу и детенышей, передними лапами, и осматривать окрестности поверх высокой травы в поисках пищи или чтобы вовремя заметить опасность. Высказывается также предположение, что общие предки прямоходящих гоминид (в том числе людей и австралопитеков) жили на мелководье, и питались мелкими водными обитателями, а прямохождение сформировалось как адаптация к передвижению по мелководью. В пользу этой версии говорит ряд анатомо-физиологических и этологических особенностей, в частности — способность людей произвольно задерживать дыхание, на что способны не все плавающие животные.
Высказывается также предположение, что общие предки прямоходящих гоминид (в том числе людей и австралопитеков) жили на мелководье, и питались мелкими водными обитателями, а прямохождение сформировалось как адаптация к передвижению по мелководью. В пользу этой версии говорит ряд анатомо-физиологических и этологических особенностей, в частности — способность людей произвольно задерживать дыхание, на что способны не все плавающие животные.
Согласно данным генетики, признаки прямохождения появились у некоторых вымерших видов обезьян ещё около 6 млн лет назад, в эпоху расхождения между людьми и шимпанзе. Это значит, что не только сами австралопитеки, но и вид, который был их предком, например, ардипитек, уже мог быть прямоходящим. Возможно, прямохождение было элементом адаптации к жизни на деревьях. Современные орангутаны используют все четыре лапы для перемещения только по толстым сучьям, в то время как к более тонким ветвям либо цепляются снизу, либо идут по ним на задних лапах, готовясь передними уцепиться за другие ветви повыше или балансируя для устойчивости. Такая тактика позволяет им приблизиться к плодам, расположенным далеко от ствола, или прыгать с одного дерева на другое. Изменения климата, произошедшие 11-12 млн лет назад, привели к сокращению лесных массивов в Африке и появлению больших открытых пространств, что и могло подтолкнуть предков австралопитеков к переходу к прямохождению по земле. В отличие от них предки современных шимпанзе и горилл специализировались на лазании по вертикальным стволам и лианам, чем обусловлена их кривоногая и косолапая походка на земле. Тем не менее, люди унаследовали много общего с этими обезьянами, включая строение костей рук, усиленных для хождения с опорой на костяшки пальцев[3][4].
Такая тактика позволяет им приблизиться к плодам, расположенным далеко от ствола, или прыгать с одного дерева на другое. Изменения климата, произошедшие 11-12 млн лет назад, привели к сокращению лесных массивов в Африке и появлению больших открытых пространств, что и могло подтолкнуть предков австралопитеков к переходу к прямохождению по земле. В отличие от них предки современных шимпанзе и горилл специализировались на лазании по вертикальным стволам и лианам, чем обусловлена их кривоногая и косолапая походка на земле. Тем не менее, люди унаследовали много общего с этими обезьянами, включая строение костей рук, усиленных для хождения с опорой на костяшки пальцев[3][4].
Не исключено также, что австралопитеки не были прямыми предками людей, а представляли собой тупиковую ветвь эволюции. К таким выводам подталкивают, в частности, недавние находки сахелантропа, ещё более древней человекообразной обезьяны, которая была больше похожа на Homo erectus, чем австралопитеки. В 2008 году был открыт новый вид австралопитеков, A. sediba, живший в Африке менее двух миллионов лет назад. Хотя по отдельным морфологическим признакам он ближе к людям, чем более древние виды австралопитеков, что дало основание его открывателям объявить его переходной формой от австралопитеков к людям, одновременно с ним, по-видимому, уже существовали первые представители рода Homo[5], такие, как человек рудольфский, что исключает возможность того, что этот вид австралопитеков мог быть предком современного человека.
В 2008 году был открыт новый вид австралопитеков, A. sediba, живший в Африке менее двух миллионов лет назад. Хотя по отдельным морфологическим признакам он ближе к людям, чем более древние виды австралопитеков, что дало основание его открывателям объявить его переходной формой от австралопитеков к людям, одновременно с ним, по-видимому, уже существовали первые представители рода Homo[5], такие, как человек рудольфский, что исключает возможность того, что этот вид австралопитеков мог быть предком современного человека.
Большинство видов австралопитеков использовали орудия труда не более современных обезьян. Известно, что шимпанзе и гориллы способны колоть орехи камнями, использовать палочки для извлечения термитов и дубинки для охоты. Насколько часто австралопитеки охотились — вопрос спорный, так как их ископаемые останки редко ассоциируют с останками убитых животных.
См. также
Примечания
Ссылки
Австралопитеки
☰
Австралопитеки – это вымершая группа семейства Гоминид, сочетавшая в себе разную степень прямохождения подобное человеческому с размером мозга чуть больше шимпанзе.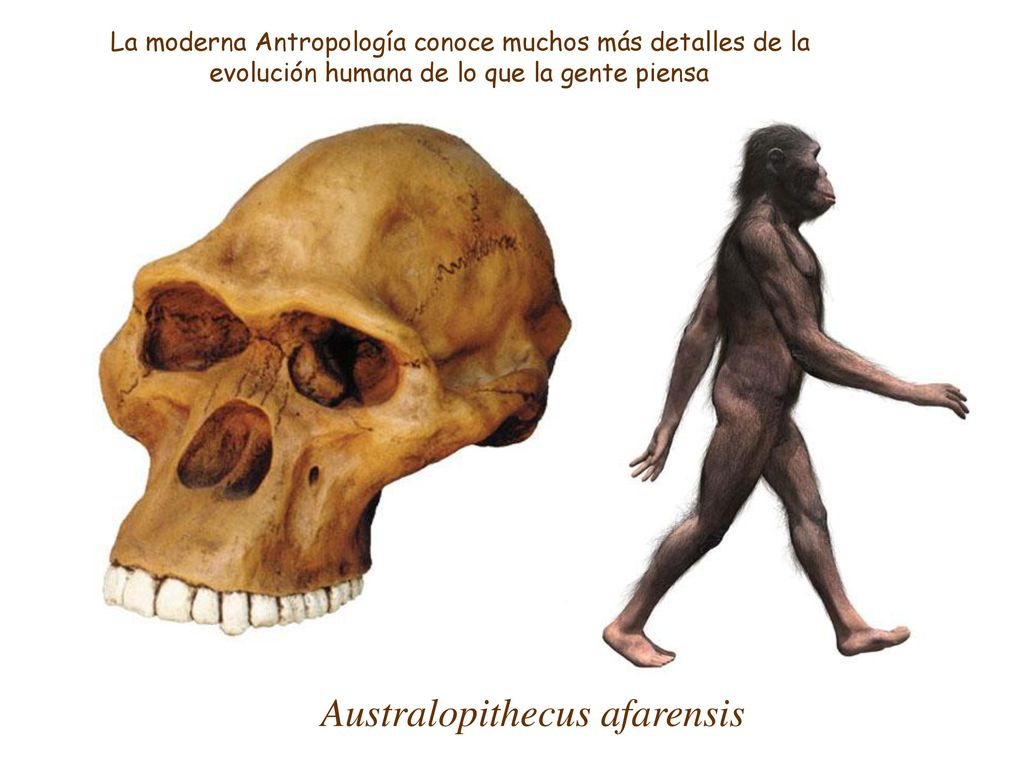
Если рассматривать всю группу, она существовала примерно с 7 до 1 миллиона лет назад. В ней выделяют роды ранних австралопитеков (до 4 миллионов лет назад), виды грацильных австралопитеков (4-2 миллиона лет назад) и род массивных австралопитеков (2.5-1 миллион лет назад). В определенном смысле истинными австралопитеками можно считать только грацильных.
Массивные австралопитеки – парантропы – сестринский роду Homo (Люди). Считается что оба рода берут начало от одного из видов грацильного австралопитека – Австралопитека Афарского. Возможно через переходные формы. Paranthropus сформировался около 2.6 миллионов лет назад, Homo – около 2.4 миллионов.
Модели: в центре – Австралопитек Афарский, слева – Парантроп Бойса, справа – Человек УмелыйТакже Австралопитек Афарский скорее всего был предком большинства других видов грацильных австралопитеков, в том числе Австралопитека Африканского.
С другой стороны, есть ряд доказательств, что предки рода Homo отделились в свою ветвь более 5 миллионов лет назад. В таком случае Австралопитек Афарский является нам родственным видом, а не прямым предком. Также некоторые находки вносят сомнение в происхождение парантропов. Их ветка также могла отделиться ранее и существовать параллельно с грациалными австралопитеками.
В таком случае Австралопитек Афарский является нам родственным видом, а не прямым предком. Также некоторые находки вносят сомнение в происхождение парантропов. Их ветка также могла отделиться ранее и существовать параллельно с грациалными австралопитеками.
Рост составлял от 1 метра до 1.5, причем самцы были на 50% крупнее самок. Объем мозга около 400-600 см3.
Обитали в Северной, Восточной и Южной Африке. Жили преимущественно в саванне.
Устройство группы напоминало таковое у шимпанзе, когда самцы кооперируются для защиты территории и самок. Из подросшего молодняка группу покидали самки. Однако для Австралопитека Афарского указывают, что мог быть один самец, владеющей группой самок.
Таз был широким, что важно для прямохождения. Большой палец хорошо развит. Челюсти слабо развиты без выступающих клыков. Австралопитеки были еще полностью покрыты шерстью, хотя вероятно уже менее густой.
Могли орудовать палками и камнями подобно современным шимпанзе. Другое дело, что прямохождение освободило руки, и делали они это чаще.
Другое дело, что прямохождение освободило руки, и делали они это чаще.
Питались в основном разнообразной растительной пищей, от листьев до фруктов и кореньев. Мясо употребляли редко.
Австралопитеки оказались «маменькиными сыночками», выяснили ученые
https://ria.ru/20190715/1556551432.html
Австралопитеки оказались «маменькиными сыночками», выяснили ученые
Австралопитеки оказались «маменькиными сыночками», выяснили ученые — РИА Новости, 15.07.2019
Австралопитеки оказались «маменькиными сыночками», выяснили ученые
Анализ зубов детеныша австралопитека показал, что наши древнейшие предки питались молоком матери так же долго, как и современные младенцы. Это объясняет то,… РИА Новости, 15.07.2019
2019-07-15T19:11
2019-07-15T19:11
2019-07-15T19:11
палеонтология
эволюция
люди
открытия — риа наука
юар
наука
/html/head/meta[@name=’og:title’]/@content
/html/head/meta[@name=’og:description’]/@content
https://cdn23. img.ria.ru/images/155654/78/1556547875_114:85:850:499_1920x0_80_0_0_893e23d1e20f0c6f14e66f4c94f03da8.jpg
img.ria.ru/images/155654/78/1556547875_114:85:850:499_1920x0_80_0_0_893e23d1e20f0c6f14e66f4c94f03da8.jpg
МОСКВА, 15 июл – РИА Новости. Анализ зубов детеныша австралопитека показал, что наши древнейшие предки питались молоком матери так же долго, как и современные младенцы. Это объясняет то, почему древних людей было так мало, и раскрывает большую роль заботы о потомстве в эволюции рода Homo, пишут ученые в журнале Nature.Как отмечает антрополог, мы почти ничего не знаем о том, как выглядели детеныши австралопитеков и первых представителей рода Homo, развивались ли они так же, как современные младенцы или были больше похожи в этом отношении на маленьких обезьян.В отличие от человека, детеныши приматов не рождаются беспомощными и очень быстро прекращаются питаться материнским молоком. Почему это так, ученые пока не знают, что постоянно порождает массу антропологических и эволюционных споров.К примеру, многие исследователи считают, что человек рождается на свет с фактически недоразвитым мозгом из-за развития прямохождения, так как узкий таз, приспособленный к движению на двух ногах, не позволяет крупному плоду уместиться в утробе матери. С другой стороны, многие исследователи предполагают, что необычный «отложенный» характер развития человеческих детей связан с метаболизмом. По их мнению, организм женщины просто не смог бы одновременно поддерживать себя и более крупный плод, если бы он продолжал развиваться внутри утробы более девяти месяцев.Фьоренца и его коллеги сделали большой шаг к разрешению всех этих споров и раскрытию роли заботы о потомстве в эволюции человека, изучая структуру и химический состав зубов двух древних австралопитеков. Их останки были открыты еще в прошлом столетии в пещере Стеркфонтейн на северо-востоке ЮАР.Для чего антропологам изучать состав зубов? Дело в том, что относительно недавно ученые обнаружили, что эмаль коренных зубов людей и животных представляет собой своеобразный «химический паспорт», отражающий в себе многие детали жизни их обладателя.К примеру, доли изотопов стронция и тяжелого кислорода-18 в эмали зубов и в костной ткани человека позволяют понять, где он родился и в каких регионах страны он жил впоследствии, а соотношение атомов кальция, бария и стронция — то, какой пищей он питался.
С другой стороны, многие исследователи предполагают, что необычный «отложенный» характер развития человеческих детей связан с метаболизмом. По их мнению, организм женщины просто не смог бы одновременно поддерживать себя и более крупный плод, если бы он продолжал развиваться внутри утробы более девяти месяцев.Фьоренца и его коллеги сделали большой шаг к разрешению всех этих споров и раскрытию роли заботы о потомстве в эволюции человека, изучая структуру и химический состав зубов двух древних австралопитеков. Их останки были открыты еще в прошлом столетии в пещере Стеркфонтейн на северо-востоке ЮАР.Для чего антропологам изучать состав зубов? Дело в том, что относительно недавно ученые обнаружили, что эмаль коренных зубов людей и животных представляет собой своеобразный «химический паспорт», отражающий в себе многие детали жизни их обладателя.К примеру, доли изотопов стронция и тяжелого кислорода-18 в эмали зубов и в костной ткани человека позволяют понять, где он родился и в каких регионах страны он жил впоследствии, а соотношение атомов кальция, бария и стронция — то, какой пищей он питался. В данном случае, ученых интересовали доли бария, так как концентрация этого элемента в молоке матери заметно выше, чем в других видах пищи. Соответственно, отслеживая его концентрацию в разных слоях эмали и дентина, можно понять, как долго их обладатель пил молоко, когда он переключился на другую пищу и пил ли он молоко впоследствии.Как оказалось, оба австралопитека, которых изучили Фьоренца и его коллеги, были «маменькиными сыночками». Они постоянно пили молоко на протяжении как минимум девяти первых месяцев своей жизни и начали переходить на другие источники пропитания лишь после первого дня рождения.Более того, ученые заметили необычные флуктуации в уровне бария, указавшие на то, что юные австралопитеки могли периодически возвращаться к грудному молоку в последующие 4-5 лет жизни. Антропологи предполагают, что подобная «сезонность» не была случайной – судя по схожим колебаниям в уровне лития, древние гоминиды, скорее всего, поступали так в тяжелые времена, когда пищи на всех не хватало.
В данном случае, ученых интересовали доли бария, так как концентрация этого элемента в молоке матери заметно выше, чем в других видах пищи. Соответственно, отслеживая его концентрацию в разных слоях эмали и дентина, можно понять, как долго их обладатель пил молоко, когда он переключился на другую пищу и пил ли он молоко впоследствии.Как оказалось, оба австралопитека, которых изучили Фьоренца и его коллеги, были «маменькиными сыночками». Они постоянно пили молоко на протяжении как минимум девяти первых месяцев своей жизни и начали переходить на другие источники пропитания лишь после первого дня рождения.Более того, ученые заметили необычные флуктуации в уровне бария, указавшие на то, что юные австралопитеки могли периодически возвращаться к грудному молоку в последующие 4-5 лет жизни. Антропологи предполагают, что подобная «сезонность» не была случайной – судя по схожим колебаниям в уровне лития, древние гоминиды, скорее всего, поступали так в тяжелые времена, когда пищи на всех не хватало. Сам факт того, что у них была возможность пить молоко матери через 4-5 лет после рождения, как считает Фьоренца, говорит о двух вещах. С одной стороны, это открытие указывает на то, что самки австралопитеков очень долго ухаживали за своим потомством и были привязаны к нему. Жизнь детенышей, по всей видимости, сильно зависела от заботы их матерей.С другой стороны, подобное поведение самок и их потомства указывает на то, что первые производили детей на свет не так часто, как это делают многие современные приматы, вкладывая все усилия в «качественное» ухаживание за немногочисленными детенышами.Это предположение хорошо укладывается в популярные современные эволюционные теории, так называемые К-стратегии, связывающие развитие рода Homo с заботой о беспомощных детенышах. Если это действительно так, то тогда группы австралопитеков и последующих гоминид должны были больше быть похожи на коллективы современных людей или семьи китов-косаток, чем на стаи других приматов, заключают ученые.
Сам факт того, что у них была возможность пить молоко матери через 4-5 лет после рождения, как считает Фьоренца, говорит о двух вещах. С одной стороны, это открытие указывает на то, что самки австралопитеков очень долго ухаживали за своим потомством и были привязаны к нему. Жизнь детенышей, по всей видимости, сильно зависела от заботы их матерей.С другой стороны, подобное поведение самок и их потомства указывает на то, что первые производили детей на свет не так часто, как это делают многие современные приматы, вкладывая все усилия в «качественное» ухаживание за немногочисленными детенышами.Это предположение хорошо укладывается в популярные современные эволюционные теории, так называемые К-стратегии, связывающие развитие рода Homo с заботой о беспомощных детенышах. Если это действительно так, то тогда группы австралопитеков и последующих гоминид должны были больше быть похожи на коллективы современных людей или семьи китов-косаток, чем на стаи других приматов, заключают ученые.
https://ria. ru/20160228/1380917545.html
ru/20160228/1380917545.html
https://ria.ru/20180706/1524111973.html
юар
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
2019
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
Новости
ru-RU
https://ria.ru/docs/about/copyright.html
https://xn--c1acbl2abdlkab1og.xn--p1ai/
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
https://cdn25.img.ria.ru/images/155654/78/1556547875_34:76:1013:810_1920x0_80_0_0_6bd61f7001573db1311994d249141e05.jpgРИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
РИА Новости
internet-group@rian. ru
ru
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
палеонтология, эволюция, люди, открытия — риа наука, юар
МОСКВА, 15 июл – РИА Новости. Анализ зубов детеныша австралопитека показал, что наши древнейшие предки питались молоком матери так же долго, как и современные младенцы. Это объясняет то, почему древних людей было так мало, и раскрывает большую роль заботы о потомстве в эволюции рода Homo, пишут ученые в журнале Nature.«Мы впервые показали, что между матерями и детенышами существовали очень прочные связи. Оказалось, что самки не только долго кормили их грудью, но и им приходилось подкармливать их молоком и в те времена, когда пищи не хватало. Это заставляет нас переосмыслить то, как мы представляем себе группы австралопитеков», — рассказывает Лука Фьоренца (Luca Fiorenza) из университета Монаша в Мельбурне (Австралия).
Как отмечает антрополог, мы почти ничего не знаем о том, как выглядели детеныши австралопитеков и первых представителей рода Homo, развивались ли они так же, как современные младенцы или были больше похожи в этом отношении на маленьких обезьян.
В отличие от человека, детеныши приматов не рождаются беспомощными и очень быстро прекращаются питаться материнским молоком. Почему это так, ученые пока не знают, что постоянно порождает массу антропологических и эволюционных споров.
28 февраля 2016, 09:57НаукаАрхеологи выяснили, что ели дети в средневековой АнглииБританские антропологи выяснили, что и богатые, и бедные дети в Англии в конце средневековья питались примерно одинаковой пищей, несмотря на разницу в статусе и достаточно быстро переставали пить молоко матери, изучив эмаль нескольких древних молочных зубов.К примеру, многие исследователи считают, что человек рождается на свет с фактически недоразвитым мозгом из-за развития прямохождения, так как узкий таз, приспособленный к движению на двух ногах, не позволяет крупному плоду уместиться в утробе матери.
С другой стороны, многие исследователи предполагают, что необычный «отложенный» характер развития человеческих детей связан с метаболизмом. По их мнению, организм женщины просто не смог бы одновременно поддерживать себя и более крупный плод, если бы он продолжал развиваться внутри утробы более девяти месяцев.
Фьоренца и его коллеги сделали большой шаг к разрешению всех этих споров и раскрытию роли заботы о потомстве в эволюции человека, изучая структуру и химический состав зубов двух древних австралопитеков. Их останки были открыты еще в прошлом столетии в пещере Стеркфонтейн на северо-востоке ЮАР.
Для чего антропологам изучать состав зубов? Дело в том, что относительно недавно ученые обнаружили, что эмаль коренных зубов людей и животных представляет собой своеобразный «химический паспорт», отражающий в себе многие детали жизни их обладателя.
К примеру, доли изотопов стронция и тяжелого кислорода-18 в эмали зубов и в костной ткани человека позволяют понять, где он родился и в каких регионах страны он жил впоследствии, а соотношение атомов кальция, бария и стронция — то, какой пищей он питался.
В данном случае, ученых интересовали доли бария, так как концентрация этого элемента в молоке матери заметно выше, чем в других видах пищи. Соответственно, отслеживая его концентрацию в разных слоях эмали и дентина, можно понять, как долго их обладатель пил молоко, когда он переключился на другую пищу и пил ли он молоко впоследствии.
Как оказалось, оба австралопитека, которых изучили Фьоренца и его коллеги, были «маменькиными сыночками». Они постоянно пили молоко на протяжении как минимум девяти первых месяцев своей жизни и начали переходить на другие источники пропитания лишь после первого дня рождения.
Более того, ученые заметили необычные флуктуации в уровне бария, указавшие на то, что юные австралопитеки могли периодически возвращаться к грудному молоку в последующие 4-5 лет жизни. Антропологи предполагают, что подобная «сезонность» не была случайной – судя по схожим колебаниям в уровне лития, древние гоминиды, скорее всего, поступали так в тяжелые времена, когда пищи на всех не хватало.
Сам факт того, что у них была возможность пить молоко матери через 4-5 лет после рождения, как считает Фьоренца, говорит о двух вещах. С одной стороны, это открытие указывает на то, что самки австралопитеков очень долго ухаживали за своим потомством и были привязаны к нему. Жизнь детенышей, по всей видимости, сильно зависела от заботы их матерей.
С другой стороны, подобное поведение самок и их потомства указывает на то, что первые производили детей на свет не так часто, как это делают многие современные приматы, вкладывая все усилия в «качественное» ухаживание за немногочисленными детенышами.
Это предположение хорошо укладывается в популярные современные эволюционные теории, так называемые К-стратегии, связывающие развитие рода Homo с заботой о беспомощных детенышах. Если это действительно так, то тогда группы австралопитеков и последующих гоминид должны были больше быть похожи на коллективы современных людей или семьи китов-косаток, чем на стаи других приматов, заключают ученые.
Кого можно считать человеком? Мы до сих пор не знаем
- Колин Баррас
- BBC Earth
Автор фото, Getty
Мы не знаем, можно ли причислить к людям неандертальцев или шимпанзе, потому что никак не можем договориться о том, какие характерные черты определяют принадлежность к роду Homo, разводит руками обозреватель BBC Earth.
Есть мнение, что человека делает человеком его культура. Часто также упоминаются душа, мораль, язык и даже чувство юмора.
Но давайте попытаемся отвлечься от метафизических категорий и разобраться в том, что делает человека человеком в фундаментально-физическом смысле.
Как это ни удивительно, общепризнанного ответа на это вопрос нет. Ученые пока так и не смогли сформулировать формальное описание нашего биологического рода Homo (люди) и нашего вида sapiens (человек разумный).
Не то чтобы ученые никогда не пытались этого сделать. На самом деле, есть сразу несколько предлагаемых возможных описаний людского рода, и в научной среде существует на удивление широкий спектр мнений по поводу того, какие черты для человека характерны, а какие — нет.
Одни специалисты считают, что род Homo существует немногим более 100 тысяч лет, и отказываются причислять к нему даже наших самых известных доисторических предков, неандертальцев.
Другие же заявляют, что человеческому роду уже примерно 11 миллионов лет, и он включает в себя не только современных людей и вымерших неандертальцев, но и шимпанзе и даже горилл.
Как получилось, что по столь фундаментальному вопросу существуют такие большие разногласия? И, в конце концов, какое из определений рода Homo является верным?
Автор фото, Alexander Roslin
Подпись к фото,Карл Линней (на портрете работы Александра Рослина, 1775 год) положил начало путанице
«Это и есть главный вопрос», — замечает Джеффри Шварц из Питтсбургского университета в американском штате Пенсильвания.
Похоже, путаница началась еще с Карла Линнея, жившего в XVIII веке шведского естествоиспытателя, который первым создал единую систему классификации растительного и животного мира.
В своей основополагающей работе «Система природы», первое издание которой вышло в 1735 году, он дал четкие названия и определения тысячам видов, но к нашему собственному роду подошел несколько более отвлеченно.
Называя очередной род животных, Линней подробно описывал его определяющие физические признаки. Но по поводу рода Homo он просто написал по-латыни «nosce te ipsum», или «познай себя».
Возможно, Линней полагал, что человек настолько сильно отличается от остальных животных, что давать ему формальное физическое описание нет необходимости.
Или же ссылался на тот факт, что люди — единственные животные, обладающие достаточным самосознанием для размышлений о собственном существовании.
В любом случае, его формулировка явно указывает на то, что люди фундаментально отличаются от всех остальных.
Линнея можно понять: он жил более чем за сто лет до публикации работ Чарльза Дарвина по теории эволюции путем естественного отбора, из которых можно сделать вывод, что люди тоже являются частью животного мира.
Но некоторые современные ученые — Шварц и его единомышленники — утверждают, что решение Линнея способно объяснить, почему человеческий род по-прежнему с трудом поддается формальному определению.
Автор фото, Thinkstock
Подпись к фото,Вот оно, секретное оружие человека — мозг!
Многие специалисты по эволюции человека вообще-то считают, что с определением рода Homo никакой проблемы нет. По их мнению, первые люди появились в Африке два-три миллиона лет назад.
До тех пор на этом континенте жили группы высших приматов, в основном относившихся к другому роду — к австралопитекам.
Австралопитеки имели ряд человеческих черт — в частности, они перемещались на двух ногах, — но их мозг был существенно меньше нашего, а передние конечности были длиннее и более приспособлены к лазанию по деревьям, как и у других обезьян. Питались они тоже иначе, чем мы.
«Принято считать, что по мере того, как их мозг постепенно увеличивался, гоминини (люди, шимпанзе и некоторые их вымершие предки – Ред.) начали употреблять в пищу мясо, и их телесные пропорции постепенно стали ближе к современному человеку, к Homo», — говорит Бернард Вуд из Университета Джорджа Вашингтона в Вашингтоне.
Но не факт, что эта распространенная теория на самом деле верна.
Самые ранние виды из обычно относимых к роду Homo имеют ряд австралопитекских черт. К примеру, человек рудольфский (Homo rudolfensis), живший примерно два миллиона лет назад, имел большое, широкое обезьяноподобное лицо, а не более узкое человеческое.
Кроме того, ранее было принято считать, что на заре нашего рода объем человеческого мозга вырос скачкообразно, но современный детальный анализ заставляет предположить, что это процесс был куда более постепенным.
Другими словами, некогда четкая граница между первыми людьми и предками-австралопитеками стала более размытой.
Автор фото, Thinkstock
Подпись к фото,Австралопитеки больше походили на обезьян
Именно этого и следовало ожидать, заявляет Брайан Вилмор из Университета Невады в американском Лас-Вегасе. По его словам, традиционно рисуемая картина ранних стадий развития человечества нуждается в корректировке.
Разглядывать окаменелые кости и пытаться понять, когда они начали в достаточной мере напоминать современные человеческие — это весьма субъективный подход.
Вместо этого определение роду Homo нужно давать, глядя на древо его эволюции.
На каком-то этапе предки человека обособились от австралопитеков, и род Homo берет свое начало именно во время этого отделения.
Сопутствующие физические характеристики, к примеру, большой мозг, появились позднее, в результате десятков тысяч лет эволюции.
Самые ранние люди были настолько близки к австралопитекам, что выглядели практически так же, считает Вилмор: имели длинные руки, маленький мозг и так далее.
На формирование собственных уникальных физических черт человечеству потребовалось не так уж много времени, но на первых порах отличить человека от австралопитека смог бы только внимательный глаз.
Вилмор таким глазом обладает. В 2015 году он и его коллеги объявили о находке, которая, по их мнению, стала самым ранним археологическим свидетельством существования рода Homo: они нашли фрагмент челюсти возрастом в 2,8 миллиона лет.
Автор фото, Getty
Подпись к фото,Неандертальцы были нашими близкими родственниками. Или все-таки нет?
Человеческой, по мнению ученых, ее можно считать благодаря нескольким малозаметным деталям. К примеру, форма небольшого отверстия в кости, через которое проходили нервы и кровеносные сосуды, однозначно человеческая, не похожая на таковую у австралопитека.
Если мы хотим составить перечень физических характеристик, определяющих род Homo, то, возможно, нам стоит обращать внимание именно на такие мелкие детали, а не на более очевидные черты вроде крупного мозга.
Но с этой точкой зрения согласны не все.
Вуд, к примеру, считает, что человеческий род появился позднее, когда у наших предков сформировался характерный образ жизни, явно отличающийся от образа жизни австралопитеков.
Последние имели длинные руки и, судя по всему, проводили немало времени на деревьях.
Человек же, как правило, живет на земле и имеет более короткие передние конечности.
Кроме того, похоже, что австралопитеки достигали зрелости сравнительно быстро, как современные обезьяны. У человека — более долгое детство.
Вуд говорит, что род Homo зародился тогда, когда наши предки наконец слезли с деревьев и продолжительность их детства начала расти.
Автор фото, Getty
Подпись к фото,Реконструкция австралопитека, получившего имя Люси
Если он прав, то при вынесении определения человеческого рода нужно учитывать и эти поведенческие черты, а не только особенности анатомии.
Применение такого подхода тоже внесет некоторые изменения в традиционное видение человеческой эволюции.
Вилмор и его коллеги не дали название виду, к которому относится найденный ими фрагмент челюсти возрастом в 2,8 миллиона лет.
Но в целом принято считать, что примерно два миллиона лет назад род Homo разделился по меньшей мере на три вида — человек умелый (H. habilis), человек рудольфский (H. rudolfensis) и человек прямоходящий (H. erectus).
Вуд полагает, что из всех трех к человеческому роду можно отнести только человека прямоходящего.
«Та отрывочная информация, которой мы обладаем о жизненном цикле habilis и rudolfensis, заставляет предположить, что они не сильно отличались от австралопитеков», — говорит он.
Кроме того, из найденных археологических фрагментов можно сделать вывод, что человек умелый сохранял способность австралопитеков ловко лазить по деревьям.
Нам нужно исключить человека умелого и человека рудольфского из нашего рода, утверждает Вуд. Их — по крайней мере, по имеющимся данным — стоит скорее отнести к австралопитекам.
Автор фото, Thinkstock
Подпись к фото,Современный человек и человек прямоходящий (Homo erectus)
У такого подхода есть недостаток: ведь в ходе изучения эволюции человека появляются новые факты, которые еще сильнее запутывают дело.
Никто не спорит с тем, что человек прямоходящий был по пропорциям своего тела близок к нам и проводил большую часть жизни на земле, а не на деревьях.
Но в 2001 году стало известно, что взрослел он гораздо быстрее, чем мы. «Его жизненный цикл существенно отличался от такового у современных людей», — говорит Вуд.
Так что же, человека прямоходящего тоже нужно исключить из рода Homo? Или же мы должны в очередной раз пересмотреть описание рода, чтобы позволить этому человеку остаться человеком?
Вуду больше по душе второй вариант, но он вызовет некоторые научные последствия: «Если включить [в наш род] человека прямоходящего, придется признать, что в этом роду есть виды с разными жизненными циклами. Эта характеристика у них различается».
Может быть, к определению человека нужно подойти вообще с другой стороны?
В конце 1990-х годов группа биологов несколько расширила рамки этой дискуссии, рассмотрев, каким образом роды определяются в отряде приматов в целом.
Автор фото, Thinkstock
Подпись к фото,Шимпанзе — это человек или нет?
Они применили критерии скорости генетических мутаций и степени генетического разнообразия в каждом роду, чтобы рассчитать, когда они появились в процессе эволюции.
Биологи выяснили, что типичный род в отряде приматов насчитывает 7-11 миллионов лет истории. Что делает род Homo, возрастом всего в 2,8 миллиона лет, большим исключением.
Ученые заявили, что имеет смысл переклассифицировать наш род таким образом, чтобы он соответствовал по возрасту другим родам приматов. То есть удлинить его историю втрое.
Другими словами, такой подход подразумевает, что род должен определяться по продолжительности его истории, а не по анатомическим или поведенческим характеристикам.
Но тогда мы получим неожиданный результат: если первые представители рода Homo жили 11 миллионов лет назад, то к ним нужно отнести не только ранних людей, но и шимпанзе, так как последние обособились от нас всего семь миллионов лет назад.
Шимпанзе, выходит, тоже люди?
Это предположение звучит, мягко говоря, спорно, но некоторые ученые видят в нем рациональное зерно.
Автор фото, Getty
Подпись к фото,По логике некоторых ученых, гориллы тоже могут оказаться людьми
В 2001 году группа генетиков решила подойти к определению рода Homo еще более широко: специалисты рассмотрели степень генетической изменчивости в нескольких родах млекопитающих.
У людей и у шимпанзе, как известно, совпадает около 99% ДНК (точные цифры несколько разнятся в зависимости от методики подсчета), ДНК гориллы тоже от них не очень сильно отличается.
Разные виды кошачьих, собачьих или медведей, обладающих подобной генетической схожестью, относились бы к одному и тому же роду, и для приматов, по идее, тоже не должно делаться исключений.
В соответствии с такой методикой, в человеческий род должны попадать не только шимпанзе, но и гориллы.
У этой идеи есть сторонники. В 2003 году Даррен Курно (сейчас работающий в Университете Нового Южного Уэльса в Австралии) совместно с покойным ныне Аланом Торном на основании исследований ДНК пытался переработать научную классификацию наших вымерших предков.
Курно и Торн указали на то, что люди и шимпанзе, несмотря на значительное сходство генов, внешне сильно отличаются друг от друга.
Автор фото, Thinkstock
Подпись к фото,Стоунхендж. Ведь это дело рук человека?
На основании этого наблюдения можно сделать вывод, что у приматов с легкостью эволюционируют новые физические черты — даже если ДНК остается почти неизменной.
Исследователи сочли, что не стоит давать названия новым ископаемым видам и родам гомининов (в отличие от «гоминини» к гомининам относят еще и горилл – Ред.) на основании одних лишь небольших различий в их костной структуре.
Все кости, похожие на человеческие и имеющие возраст до семи миллионов лет, должны относиться к роду Homo, заявили они. А заодно в род нужно включить и шимпанзе.
Однако сейчас Курно говорит, что с тех пор изменил свою точку зрения. «Я больше не считаю, что шимпанзе нужно относить к роду Homo», — говорит ученый.
Теперь Курно скорее согласен с теорией, предлагаемой Вилмором и его единомышленниками, а именно: люди впервые появились около 2,8 млн лет назад и были тогда представлены такими видами, как Homo habilis и Homo gautengensis (этот вид Курно описал на основании окаменелостей, найденных в Южной Африке).
Хотя Курно сейчас и не согласен с собственными ранними выводами, они, по крайней мере, представляли собой попытку привести определение рода Homo в соответствие с принятыми практиками классификации других приматов и млекопитающих — а заодно уточнить расплывчатое определение, сформулированное Линнеем 280 лет назад.
Вуд тоже говорит, что он преследует именно эту цель в своих поисках описания рода человека.
Автор фото, B Christopher Alamy Stock Photo
Подпись к фото,Homo florensiensis — человек или нет?
Шварц тоже хотел бы сформулировать определение Homo таким образом, чтобы оно не выделялось из системы классификации других родов млекопитающих.
Но результат — если последовать его методике — окажется совершенно иным.
Шварц считает, что самые главные отличия между родами млекопитающих заключаются во внешности, а не в генах и не в поведении.
«Выдры вскрывают ракушки при помощи камней, вороны умеют с помощью камешков поднять уровень воды в сосуде, чтобы до нее дотянуться, но мы ведь не станем утверждать, что эти особенности поведения определяют выдр или ворон, — замечает он. — И к гомининам нужно относиться точно так же, как и к любым другим организмам».
По его словам, если внимательно присмотреться к ископаемым останкам гомининов, в них обнаружится гораздо больше разнообразия, чем считает большинство исследователей.
К примеру, неандертальцы были крепче сбиты, чем мы, и имели выраженные надбровные дуги, которых, как правило, нет у современных людей.
Шварц утверждает, что если бы подобные различия наблюдались у других животных, ученые без колебаний отнесли бы их к совершенно разным родам.
Тут уж не до шимпанзе: Шварц предлагает хорошенько задуматься над тем, не изгнать ли из наших сородичей даже неандертальцев.
По его словам, нужно начать с известной величины — ныне живущих людей — и постепенно двигаться назад во времени, внимательно оценивая, какие окаменелости действительно относятся к нашему роду и виду.
«Эта точка зрения не очень популярна, но если мы хотим классифицировать человека точно так же, как свиней, грызунов, лошадей и других млекопитающих, именно так нам и нужно поступить», — убежден ученый.
Автор фото, SLP
Подпись к фото,Можно ли принять в нашу семью Homo erectus?
Он уже начал работать по этой методике, обращая особое внимание на черепа и челюсти гомининов.
Человеческий род определяется формой подбородка и лба, говорит он. А их характерная форма впервые появилась примерно 100 тысяч лет назад.
Если исходить из этого, то родственниками ныне живущих людей можно считать лишь относительно недавних наших предков — такие останки были найдены, к примеру, в израильской пещере Схул и в южноафриканской пещере Бордер-кейв.
В общем, совершенно ясно, что недостатка в разных научных методологиях определения нашего рода нет.
Но у ученых также нет и консенсуса по поводу того, какое из определений нужно считать правильным. А с учетом того, насколько рьяно сторонники разных точек зрения отстаивают свои позиции, ожидать компромисса в ближайшем будущем вряд ли приходится.
Может показаться удивительным, что мы не способны без споров определить саму нашу суть. Но, возможно, дело тут отчасти и в том, что речь идет о человечестве.
«Когда мы сравниваем окаменелые кости лошадей, страсти не кипят, — замечает Шварц. — А если речь идет о гомининах, начинают бушевать эмоции».
австралопитеков | Характеристики и факты
Идентификация самого раннего представителя человеческого племени (Hominini) затруднительна, потому что предки современных людей становятся все более обезьяноподобными по мере того, как летопись окаменелостей прослеживается во времени. Они напоминают то, что можно было бы ожидать от общего предка людей и обезьян, тем, что они обладают сочетанием человеческих и обезьяньих черт. Например, предполагаемый самый ранний вид, Sahelanthropus tchadensis , похож на человека тем, что имеет слегка уменьшенный клык и лицо, которое не выступает далеко вперед.Однако во многих других отношениях, включая размер мозга, он похож на обезьяну. Неизвестно, ходили ли эти животные прямо, потому что были найдены только один череп, фрагменты одной или нескольких нижних челюстей и несколько зубов.
Получите подписку Britannica Premium и получите доступ к эксклюзивному контенту. Подпишитесь сейчасДвуногие, однако, по-видимому, установились у 6 миллионов лет назад Orrorin tugenensis , австралопитека до , найденного на холмах Туген возле озера Баринго в центральной Кении.В 2001 году эти окаменелости были описаны как самые ранние из известных гомининов. O. tugenensis является примитивным по большей части, если не по всей своей анатомии, за исключением бедренных костей (бедренные кости), которые, по-видимому, имеют общие черты двуногости с современными людьми. Как и более поздние гоминины, у него есть зубы с толстой коренной эмалью, но, в отличие от людей, у него отчетливо обезьяноподобные клыки и премоляры. В основе его статуса гоминина лежат человеческие особенности бедренной кости. По словам его первооткрывателей, особенности бедренной кости, подразумевающие двуногие, включают ее общие пропорции, внутреннюю структуру шейки бедренной кости (столб, соединяющий шарообразную головку бедренной кости с стержнем кости) и бороздку на кости для мышца, используемая при вертикальной ходьбе (наружная запирательная мышца).
Ardipithecus kadabba и Ar. рамидусДругой кандидат на звание самого раннего гоминина относится к роду Ardipithecus (5,8–4,4 млн лет назад). Остатки ар. Кадабба (5,8–5,2 млн лет назад), обнаруженные в средней долине реки Аваш в Афарском регионе Эфиопии (впадина, расположенная в северной части страны, простирающейся на северо-восток до Красного моря), содержат фрагменты костей конечностей, изолированные зубы, частичная нижняя челюсть и кость пальца.Хотя клык в некоторых отношениях похож на обезьяну, он не демонстрирует классического взаимосвязанного хонинговального комплекса (когда внутренняя сторона верхнего клыка заостряется против нижнего премоляра [или двустворчатого клапана]). Кость пальца, относящаяся к Ardipithecus , демонстрирует двуногую анатомию, но она была обнаружена в отложениях на 400000 лет моложе и примерно в 20 км (12,4 миль) от окаменелости, использованной для определения Ar. kadabba и может принадлежать к другому виду ранних гомининов.
Ар.ramidus , который был обнаружен в средней части долины Аваш в 1992 году на стоянке под названием Арамис, известен по раздробленному и искаженному частичному скелету. Череп обезьяноподобный с крошечным мозгом — 300–350 куб. См (18,3–21,4 кубических дюйма), что эквивалентно весу мозга около 300–350 граммов (10,6–12,3 унции), и прогнатической (выступающей) мордой. Большое отверстие (большое отверстие) в основании черепа располагается под черепной коробкой, как у двуногого, а не сзади, как у четвероногой (четвероногой) обезьяны ( см. череп).
Некоторые другие анатомические характеристики Ar. ramidus предполагают, что он был приспособлен к древесным условиям. Верхняя конечность отличается от таковой у современного человека. Он очень длинный, что позволяло кончикам пальцев доходить как минимум до колена. Чрезвычайно большие руки этого вида указывают на образ жизни, который включал в себя значительные восхождения и другие занятия среди деревьев. Таз — это смесь черт человека и обезьяны; он кажется шире, короче и уже, чем таз обезьяны, и напоминает двуногий таз.Стопа особенно похожа на обезьяну, с удлиненными пальцами и полностью расходящимся большим пальцем для передвижения по деревьям. Окаменелости животных, пыльца и другие свидетельства, связанные с Ar. ramidus также указывает на то, что он был дома в лесной среде ( см. также Ardi).
Australopithecus anamensisСамый ранний представитель рода Australopithecus — Au. anamensis , который был обнаружен на севере Кении недалеко от озера Туркана в Канапой и заливе Аллия.Впервые этот вид был описан в 1995 году после анализа изолированных зубов, верхней и нижней челюстей, фрагментов черепа и большеберцовой кости, обнаруженных в местах обнаружения. Окаменелости датируются 4,2–3,9 млн лет назад и, как и Ardipithecus , Au. anamensis также ассоциируется с лесными животными и несколькими видами пастбищ.
Морда прогнатическая. Зубы имеют толстую эмаль, как и у всех более поздних гомининов, но в отличие от зубов Ar. ramidus с тонкой обезьяньей эмалью.Большеберцовая кость (большеберцовая кость) демонстрирует анатомию на концах колена и лодыжки, характерную для более поздних двуногих гомининов.
Сильно раздавленный и деформированный череп, обнаруженный в Ломекви на западном берегу озера Туркана на севере Кении в 1998 году, был отнесен к новому роду и виду, «человек из Кении», Kenyanthropus platyops (3,5 млн лет назад). Это тоже связано с лесной фауной. Является ли этот единственный экземпляр действительно новым видом, широко обсуждается, поскольку череп может быть сильно искаженным примером другого вида, Au.afarensis .
копия черепа Kenyanthropus platyopsРеплика черепа Kenyanthropus platyops возрастом от 3,2 до 3,5 миллионов лет, найденного антропологом Мивом Лики в 1998 году в Ломекви, недалеко от озера Туркана, Кения.
© Bone Clones, www.boneclones.comЭволюционная история австралопитов | Evolution: Education and Outreach
Arambourg C, Coppens Y. Sur la decouverte, dans la Pleistocene inferieur de la vallee de l’Omo [Эфиопия], d’une mandibule d’australopiecien.CR Acad Sci Paris. 1967; 265: 589–90.
CAS Google Scholar
Asfaw B, White T, Lovejoy O, Latimer B, Simpson S, Suwa G. Australopithecus garhi: новый вид ранних гоминидов из Эфиопии. Наука. 1999. 284: 629–35.
Артикул CAS Google Scholar
Berger LR, de Ruiter DJ, Churchill SE, Schmid P, Carlson KJ, Dirks PHGM и др. Australopithecus sediba: новый вид Homo, похожий на австралопит из Южной Африки. Наука. 2010. 328: 195–204.
Артикул CAS Google Scholar
Буль М. Окаменелости домов лесов: элементы человеческой палеонтологии. Париж: Массон; 1921.
Google Scholar
Брум Р. Плейстоценовые человекообразные обезьяны Южной Африки. Природа. 1938; 142: 377–9.
Артикул Google Scholar
Брум Р. Другой вид ископаемого обезьяно-человека. Природа. 1949; 163: 57.
Артикул Google Scholar
Брум Р., Робинсон Дж. Новый тип ископаемого человека. Природа. 1949; 164: 322–3.
Артикул CAS Google Scholar
Брум Р., Робинсон Дж. Еще одно свидетельство строения обезьяно-человека Стеркфонтейна Plesianthropus .Mem Transv Mus. 1950; 4: 7–84.
Google Scholar
Broom R, Schepers GWH. Южноафриканские ископаемые обезьяно-люди: Australopithecinae. Mem Trans Mus. 1946; 2: 1–272.
Google Scholar
Брюне М, Бовилан А, Коппенс Y, Хайнц Э, Мутай АХЕ, Пилбим ДР. Australopithecus bahrelghazali, une nouvelle espece d’Hominide ancien de la региона Коро-Торо.C R Acad Sci Paris. 1996; 322: 907–13.
Google Scholar
Брюнет М., Гай Ф., Пилбим Д., Маккей Х.Т., Ликиус А., Ахунта Д. и др. Новый гоминид из верхнего миоцена Чада, Центральная Африка. Природа. 2002; 418: 145–51.
Артикул CAS Google Scholar
Кларк Р.Дж. Недавно обнаруженная информация о скелете Sterkfontein Member 2 Australopithecus .S Afr J Sci. 2002; 98: 523–6.
Google Scholar
Константино П., Вуд Б. Эволюция Zinjanthropus boisei . Evol Anthropol. 2007; 16: 49–62.
Артикул Google Scholar
Дарт РА. Australopithecus africanus: человек-обезьяна из Южной Африки. Природа. 1925; 115: 195–9.
Артикул Google Scholar
Дарт РА.Проточеловек Макапансгата Australopithecus prometheus . Am J Phys Anthropol. 1948; 6: 259–83.
Артикул CAS Google Scholar
de Heinzelin J, Clark JD, White T, Hart W., Renne P, Woldegabriel G, et al. Окружающая среда и поведение гоминидов Бури возрастом 2,5 миллиона лет. Наука. 1999. 284: 625–9.
Артикул Google Scholar
Галик К., Сенут Б., Пикфорд М., Гоммери Д., Трейл Дж., Куперавадж А. и др.Внешняя и внутренняя морфология бедренной кости BAR 1002’00 Orrorin tugenensis . Наука. 2004; 305: 1450–3.
Артикул CAS Google Scholar
Gordon AD, Green DJ, Richmond BG. Сильный посткраниальный диморфизм размера у Australopithecus afarensis: является результатом двух новых методов повторной выборки для многомерных наборов данных с отсутствующими данными. Am J Phys Anthropol. 2008; 135: 311–28.
Артикул Google Scholar
Green DJ, Gordon AD, Richmond BG.Размеры конечностей Australopithecus afarensis и Australopithecus africanus . J Hum Evol. 2007. 52: 187–200.
Артикул Google Scholar
Грине ИП, Унгар ПС, Теафорд МФ. Был ли ранний плиоцен hominin ‘ Australopithecus ‘ anamensis кормушкой для твердых предметов? S Afr J Sci. 2006a; 102: 301–10.
Google Scholar
Grine FE, Ungar PS, Teaford MF, El-Zaatari S.Молярный микроножок у Praeanthropus afarensis : свидетельство диетического застоя во времени и в различных палеоэкологических условиях. J Hum Evol. 2006b; 51: 297–319.
Артикул Google Scholar
Хайле-Селассие Y, Сува G, Уайт ТД. Зубы позднего миоцена из Среднего Аваша, Эфиопия, и эволюция зубов раннего гоминида. Наука. 2004; 303: 1503–5.
Артикул CAS Google Scholar
Хауэлл ФК.Обзор плиоцена и раннего плейстоцена нижней части бассейна реки Омо, южная Эфиопия. В: Jolly C, редактор. Ранние гоминиды Африки. Лондон: Дакворт; 1978. стр. 85–130.
Google Scholar
Johanson DC, белый TD. Систематическая оценка ранних африканских гоминидов. Наука. 1979; 202: 321–30.
Артикул Google Scholar
Веселый СиДжей. Семеноеды — новая модель дифференциации гоминидов, основанная на аналогии с павианом.Мужчина. 1970; 5: 5–26.
Артикул Google Scholar
Jungers WL. Конечности Люси: аллометрия скелета и движения в Australopithecus afarensis . Природа. 1982; 297: 676–8.
Артикул Google Scholar
Кейт А. Древность человека. Лондон: Уильямс и Норгейт; 1915.
Google Scholar
Кейт А.Ископаемая человекообразная обезьяна из Таунга. Природа. 1925; 115: 234–5.
Артикул Google Scholar
Kimbel WH, Delezene LK. Редукс «Люси»: обзор исследований Australopithecus afarensis . Am J Phys Anthropol. 2009. 140: 2–48.
Артикул Google Scholar
Kimbel WH, Lockwood CA, Ward CV, Leakey MG, Rak Y, Johanson DC. Был Australopithecus anamensis предком A.afarensis ? Случай анагенеза в летописи окаменелостей гомининов. J Hum Evol. 2006; 51: 134–52.
Артикул Google Scholar
Кляйн Р.Г. Причины «стойкого» вымирания австралопитеков. В: Грине Ф.Е., редактор. Эволюционная история «крепких» австралопитеков. Нью-Йорк: Альдин де Грюйтер; 1988. с. 499–505.
Google Scholar
Куллмер О., Сандрок О., Абель Р., Шренк Ф., Бромаж Т.Г., Джувейи Ю.М.Первый Paranthropus из разлома Малави. J Hum Evol. 1999; 37: 121–7.
Артикул CAS Google Scholar
Lawn BR, Lee JJW. Анализ режимов разрушения и деформации зубов, подвергшихся окклюзионной нагрузке. Acta Biomater. 2009; 5: 2213–21.
Артикул Google Scholar
Лики Л. Новый ископаемый череп из Олдувая. Природа. 1959; 184: 491–3.
Артикул Google Scholar
Leakey MD, Hay R. Следы плиоцена в слоях Laetolil в Laetoli, северная Танзания. Природа. 1979; 232: 308–12.
Артикул Google Scholar
Leakey REF, Walker A. Australopithecus, Homo и гипотеза одного вида. Природа. 1976; 261: 572–4.
Артикул CAS Google Scholar
Лики LSB, Эвернден Дж. Ф., Кертис Г. Х.Эпоха кровати I, Олдувайское ущелье, Танганьика. Природа. 1961; 189: 649–50.
Артикул CAS Google Scholar
Leakey MG, Feibel CS, McDougall I, Ward C, Walker AC. Новые виды гоминидов возрастом четыре миллиона лет из Канапой и Аллиа Бэй, Кения. Природа. 1995; 376: 565–71.
Артикул CAS Google Scholar
Лики М.Г., Спур Ф., Браун Ф.Х., Гатого П.Н., Киари С., Лики Л.Н. и др.Новый род гоминидов из Восточной Африки показывает разнообразные линии среднего плиоцена. Природа. 2001; 410: 433–40.
Артикул CAS Google Scholar
Локвуд Калифорния, Тобиас П.В. Морфология и сходство новых черепных останков гомининов из пачки 4 формации Стеркфонтейн, провинция Гаутенг, Южная Африка. J Hum Evol. 2002; 42: 389–450.
Артикул Google Scholar
Lovejoy CO.Происхождение человека. Наука. 1981; 211: 341–50.
Артикул CAS Google Scholar
Lovejoy CO. Эволюция ходьбы человека. Sci Am. 1988. 256: 118–25.
Артикул Google Scholar
Lovejoy CO, Heiple KG, Burstein AH. Походка австралопитека . Am J Phys Anthropol. 1973; 38: 757–80.
Артикул CAS Google Scholar
Лавджой, Колорадо, Латимер Б, Сува Г, Асфау Б, Уайт ТД.Сочетание хватки и движения: стопа Ardipithecus ramidus . Наука. 2009a; 326: 72e1–8.
Google Scholar
Лавджой Колорадо, Симпсон ЮЗ, Уайт ТД, Асфау Б., Сува Г. Осторожное лазание в миоцене: передние конечности Ardipithecus ramidus и люди примитивны. Наука. 2009b; 326: 70e1–8.
Google Scholar
Лавджой, Колорадо, Сува Г, Сперлок Л, Асфау Б, Уайт ТД.Таз и бедро Ardipithecus ramidus: Возникновение прямой ходьбы. Наука. 2009c; 326: 71e1–6.
Google Scholar
Marshall AJ, Wrangham RW. Эволюционные последствия использования запасных продуктов. Int J Primatol. 2007. 28: 1218–35.
Артикул Google Scholar
Martin LB. Значение толщины эмали в эволюции гоминоидов. Природа.1985; 314: 260–3.
Артикул CAS Google Scholar
МакГенри Х.М. Относительный размер щечных зубов у австралопитека . Am J Phys Anthropol. 1984. 64: 297–306.
Артикул CAS Google Scholar
McHenry HM, Berger L. Пропорции тела Australopithecus afarensis и происхождение рода Homo . J Hum Evol.1998; 35: 1–22.
Артикул CAS Google Scholar
Олейничак А.Дж., Смит Т.М., Скиннер М.М., Грин Ф.И., Фини Р.Н.М., Теккерей Дж.Ф. и др. Трехмерное распределение и толщина моляров эмали у австралопитека и парантропа . Письма Биол. 2008; 4: 406–10.
Артикул CAS Google Scholar
Партридж TC, Granger DE, Caffee MW, Clarke RJ.Останки гоминидов нижнего плиоцена из Стеркфонтейна. Наука. 2003; 300: 607–12.
Артикул CAS Google Scholar
Петерс ЧР. Орехоподобные масличные семена — пища для обезьян, шимпанзе, людей и, вероятно, обезьянолюдей. Am J Phys Anthropol. 1987. 73: 333–63.
Артикул CAS Google Scholar
Плавкан ЖМ. Половой диморфизм в эволюции приматов. Yrbk Phys Anthropol.2001; 44: 25–53.
Артикул Google Scholar
Плавкан ЖМ. Масштабирование отношений между черепно-лицевым половым диморфизмом и диморфизмом массы тела у приматов: значение для летописи окаменелостей. Am J Phys Anthropol. 2003; 120: 38–60.
Артикул Google Scholar
Поттс Р. Экологические гипотезы эволюции гомининов. Yrbk Phys Anthropol. 1998. 41: 93–136.
Артикул Google Scholar
Рак Ю. Лицо австралопитека. Нью-Йорк: академический; 1983.
Книга. Google Scholar
Рид KE. Ранняя эволюция гоминидов и экологические изменения в африканском плио-плейстоцене. J Hum Evol. 1997. 32: 289–322.
Артикул CAS Google Scholar
Reno PL, Meindl RS, McCollum MA, Lovejoy CO.Половой диморфизм у Australopithecus afarensis был подобен таковому у современных людей. Proc Natl Acad Sci U S. A. 2003; 100: 9404–9.
Артикул CAS Google Scholar
Ричмонд, Б.Г. Биомеханика искривления фаланг. J Hum Evol. 2007; 53: 678–90.
Артикул Google Scholar
Ричмонд Б.Г., Юнгерс В.Л. Orrorin tugenensis Морфология бедренной кости и эволюция двуногости гомининов.Наука. 2008; 319: 1662–5.
Артикул CAS Google Scholar
Scott RS, Ungar PS, Bergstrom TS, Brown CA, Grine FE, Teaford MF и др. Анализ текстуры стоматологического микронного носителя показывает внутривидовую изменчивость рациона ископаемых гомининов. Природа. 2005; 436: 693–5.
Артикул CAS Google Scholar
Сенут Б., Пикфорд М., Гоммери Д., Майн П., Чебой К., Коппенс Ю.Первый гоминид из миоцена (формация Люкейно, Кения). C R Acad Sci Paris Sci Terre Plan. 2001; 332: 137–44.
Артикул Google Scholar
Smith GE. Ископаемая человекообразная обезьяна из Таунга. Природа. 1925; 115: 235.
Google Scholar
Спенсер Ф. Пилтдаун: научная подделка. Оксфорд: издательство Оксфордского университета; 1990.
Google Scholar
Sponheimer M, Passey BH, de Ruiter DJ, Guatelli-Steinberg D, Cerling TE, Lee-Thorp JA.Изотопные данные о вариабельности питания ранних гомининов Paranthropus robustus . Наука. 2006; 314: 980–2.
Артикул CAS Google Scholar
Stern JT, Susman RL. Анатомия опорно-двигательного аппарата Australopithecus afarensis . Am J Phys Anthropol. 1983; 60: 279–317.
Артикул Google Scholar
Strait DS, Grine FE. Кладистика и ранняя филогения гоминидов.Наука. 1999; 285: 1210.
Артикул CAS Google Scholar
Strait DS, Grine FE. Вывод гоминоидов и ранней филогении гоминидов с использованием краниодентальных признаков: роль ископаемых таксонов. J Hum Evol. 2004. 47: 399–452.
Артикул Google Scholar
Strait DS, Grine FE, Fleagle JG. Анализ филогении гоминидов. В: Хенке В., Таттерсолл И., редакторы. Справочник по палеоантропологии, т.3. Берлин: Спрингер; 2007. с. 1781–806.
Глава Google Scholar
Strait DS, Weber GW, Neubauer S, Chalk J, Richmond BG, Lucas PW и др. Биомеханика питания и экология питания Australopithecus africanus . Proc Natl Acad Sci U S. A. 2009; 106: 2124–9.
Артикул CAS Google Scholar
Susman RL. Новые посткраниальные останки Сварткранса и их влияние на функциональную морфологию и поведение Paranthropus robustus .В: Грине Ф.Е., редактор. Эволюционная история «крепких» австралопитеков. Нью-Йорк: Альдин де Грюйтер; 1988a. п. 149–72.
Google Scholar
Susman RL. Рука Paranthropus robustus из пачки 1, Сварткранс: ископаемые свидетельства поведения инструмента. Наука. 1988b; 240: 781–4.
Артикул CAS Google Scholar
Susman RL. Ископаемые свидетельства использования орудий раннего гоминида.Наука. 1994; 265: 1570–3.
Артикул CAS Google Scholar
Сува Джи, Уайт ТД, Хауэлл ФК. Постклыковые зубные ряды нижней челюсти из формации Шунгура, Эфиопия: морфология коронки, таксономическое распределение и эволюция плиоплейстоценовых гоминидов. Am J Phys Anthropol. 1996; 101: 247–82.
Артикул CAS Google Scholar
Тринкаус Э., Лонг Дж. Видовая принадлежность первых пястных костей пачки Сварткранса 1: SK 84 и SKX 5020.Am J Phys Anthropol. 1990; 83: 419–24.
Артикул CAS Google Scholar
Унгар ПС, Грине ИП, Теафорд МФ. Стоматологическая микропитка и диета плио-плейстоценового гоминина Paranthropus boisei . PLoS ONE. 2008; 3: e2044.
Артикул Google Scholar
Врба ЭС. О связи палеоклимата и эволюции. В: Vrba ES, Denton GH, Partridge TC, Burckle LH, редакторы.Палеоклимат и эволюция с акцентом на происхождение человека. Нью-Хейвен: издательство Йельского университета; 1995. стр. 24–45.
Google Scholar
Walker A, Leakey REF, Harris J, Brown F. 2,5 млн лет назад Australopithecus boisei к западу от озера Туркана, Кения. Природа. 1986; 322: 517–22.
Артикул Google Scholar
Уокер Дж., Клифф Р.А., Латам АГ. U – Pb изотопный возраст гоминида StW 573 из Стеркфонтейна, Южная Африка.Наука. 2006; 314: 1592–4.
Артикул CAS Google Scholar
Walter RC, Manega PC, Hay RL, Drake RE, Curtis GH. Лазерный синтез 40 Ar / 39 Ar датирование пласта I, Олдувайское ущелье, Танзания. Природа. 1991; 354: 145–9.
Артикул CAS Google Scholar
Уошберн SL. Инструменты и эволюция человека. Sci Am. 1960; 203: 63–75.
Артикул CAS Google Scholar
Белый TD.Ранние гоминиды — разнообразие или искажение? Наука. 2003; 299: 1994–7.
Артикул CAS Google Scholar
White TD, Suwa G, Asfaw B. Australopithecus ramidus, новый вид ранних гоминидов из Арамиса, Эфиопия. Природа. 1994; 371: 306–12.
Артикул CAS Google Scholar
White TD, WoldeGabriel G, Asfaw B, Ambrose S, Beyene Y, Bernor RL, et al.Аса Исси, Арамис и происхождение австралопитека . Природа. 2006; 440: 883–9.
Артикул CAS Google Scholar
White TD, Asfaw B, Beyene Y, Haile-Selassie Y, Lovejoy CO, Suwa G, et al. Ardipithecus ramidus и палеобиология ранних гоминидов. Наука. 2009. 326: 75–86.
CAS Google Scholar
Вуд BA, пролив DS.Модели использования ресурсов в начале г. Homo и Paranthropus . J Hum Evol. 2004. 46: 119–62.
Артикул Google Scholar
Род Australopithecus
Род AustralopithecusРод Australopithecus
Около 4.4 миллиона
много лет назад появился другой тип приматов.
Первые двуногие приматы классифицированы палеонтологами
как
гоминидов, и эти первые гоминиды еще не развили
большой
мозг, структура зубов и особенности скелета идентифицированы как Homo.
Вместо этого они предшествуют, а иногда и перекрывают первые виды Homo
и известны как австралопитеки.Два типа австралопитеков
— изящные и крепкие.
АНАМЕНЗ АВСТРАЛОПИТЕКА
4,0–3,9 миллиона лет назад
Этот вид недавно был обнаружен в районе озера Туркана в Кении. и восходит к 4 миллионам лет назад.Хотя не признан таковым для 30 лет назад в Канапой произошло первое открытие Australopithecus anamensis. регион Восточного озера Туркана в 1965 году экспедицией Гарвардского университета. В Первоначальная находка состояла из частичной левой плечевой кости. Помимо одиночного моляра Открытие в 1982 г., больше экземпляров A. anamensis обнаружено не было. до начала 1990-х годов, когда Мив Лики и другие дочерние компании Национальные музеи Кении организовали исследовательскую группу для региона Канапои.
Около 3,9 миллиона лет назад A. anamensis превратились в австралопитека. афарский. Это первое свидетельство окаменелостей как первое и самый ранний двуногий. Большеберцовая кость Australopithecus anamensis указывает на двуногость. Это первый вид, который может ходить прямо!
Некоторые характеристики:
Объем черепа Australopithecus anamensis неизвестен.Рост самцов около 5 футов, а у самок около 4 футов 3 дюйма. Мужчина
вес составляет около 110 фунтов, в то время как вес женщины составляет около 70 фунтов.
Их зубы и челюсти гоминидные, но имеют некоторое сходство с шимпанзе.
Они сохранили обезьяноподобный череп и зубной ряд, но при этом
развитая посткрания, более или менее типичная по форме гоминид. Суставы
кости ног указывают на двуногую походку.
http: // www.archaeologyinfo.com/australopithecusanamensis.htm
http://www.archaeologyinfo.com/ images / ramidus.JPG
АВСТРАЛОПИТЕК АФАРЕНСИС
Австралопитек афарский
Australopithecus afarensis был назван как вид в 1978 году Д.Джонхансон
и Т. Уайт. Это один из наиболее известных австралопитеков, потому что
большое количество окаменелостей было найдено и отнесено к этому виду.
Фрагменты более 300 особей Australopithecus afarensis были обнаружены до сих пор в Танзании, Кении и Эфиопии. Австралопитек
afarensis жил примерно от 4,1 до 2,7 миллиона лет назад на северо-востоке
Африка.
Самый известный экземпляр — «Люси»,
почти полный скелет, найденный в 1974 году в Хадаре, Эфиопия.Иллюстрация
справа показывает «Люси» в сравнении с современной человеческой женщиной. Она была
всего около 3 футов 8 дюймов в высоту. Самцы были несколько выше и вдвое больше.
робастный (половой диморфизм).
A. afarensis все еще очень обезьяноподобный, но имеет некоторые характеристики Homo sapiens . Например, общий размер тела Люси, размер мозга и форма черепа. напоминают шимпанзе.Однако A. afarensis ходил прямо, как человек, не как шимпанзе. Доказательства двуногости исходят от окаменелостей скелетов показывает сочленение таза и бедра (бедренную кость), аналогичные человеческим. Неопровержимый Доказательства исходят из следов Лаэтоли. Нет связанного с производством инструмента с A. afarensis.
Скелет Люси и Люси против современного человека женский
Отличительные характеристики А.afarensis были:
- низкий лоб
- костный гребень над глазами
- плоский нос и отсутствие подбородка
- более человекоподобные зубы,
- и кости таза и ног напоминали таковые у современного человека.
- Самки были меньше самцов.
Многие ученые считают, что Australopithecus afarensis все еще существовал.
активен на деревьях, потому что пальцы рук и ног этого вида были
изогнутые и длиннее, чем у современного человека.Кроме того, его верхняя часть туловища
сильнее нашего, и его длина рук и ног схожа, что также
указывает на то, что вид приспособился к лазанию по деревьям.
Следы Лаэтоли
Следы А. afarensis были обнаружены Мэри Лики в 1974 году в Лаэтоли, Танзания.Находка датируется 3,6 миллиона лет назад. Отпечатки были сделаны после вулканического извержения и были быстро покрыты последующим извержением где сохранились следы. По крайней мере, два человека ходят бок о бок представлены. Мало того, что отпечатки показывают, что A. afarensis ходили прямо, но у них также не было расходящегося пальца, как у шимпанзе.
___________________________________________________Ссылки:
Австралопитек афарский
Хронология видов
Зал предков человека: Australopithecus afarensis
Истоки человечества — Исследовательский центр: А.афарский
AUSTRALOPITHECUS BAHRELGHAZALI
3.5 — 3,0 миллиона много лет назад.
А. Бахрелгазали был обнаружен Мишелем Брюне в 1993 году в старинном
русло реки Бахр-эль-Газаль в Чаде, расположенное в 2500 км к западу от
Восточноафриканская рифтовая долина. На данный момент это первый и единственный австралопитек.
экземпляр, который можно найти в Северной и Центральной Африке, а также на самом дальнем западе
любого образца, найденного на сегодняшний день.
Существуют аргументы против того, что A. bahrelghazali является его
собственный вид из-за его сходства с Australopithecus afarensis . Однако нижнечелюстной симфиз более современен по внешнему виду.
чем у A afarensis .Хотя видовое обозначение Australopithecus bahrelghazali — новый, в настоящее время набирает
все больше и больше поддержки среди ученых.
http://www.msu.edu/~heslipst/contents/ANP440/bahrelghazali.htm
ИЗОБРАЖЕНИЕ:
http: // www.scienceinafrica.co.za/ pics / origin5a.gif
АВСТРАЛОПИТЕК АФРИКАНСКИЙ ЯЗЫК
3.0-2,5 миллиона лет назад
Раймонд Дарт открыл первого австралопитека в ноябре 1924 года.
ископаемое было найдено в известковом карьере в Таунге, к юго-западу от Йоханнесбурга, и
принадлежал к незрелой обезьяноподобной особи. Окаменелость существовала на лице, части
черепа, полной нижней челюсти и эндокаста головного мозга, образовавшегося при
песок внутри черепа превратился в камень, фиксируя форму мозга.
Дарт заявил, что таунг человек был более ранней формой человека и назвал его австралопитеком. africanus («южная обезьяна из Африка »). Australopithecus africanus оказался обезьяноподобным, имея выступающее лицо и маленький мозг, но с явно неприглядными зубными рядами, включая маленькие клыки и большие плоские коренные зубы.Двуногая поза снова стала обозначается центральным положением большого затылочного отверстия и анатомией позвоночника, таза и бедра.
http: // www.geocities.com/palaeoanthropology/Aafricanus.html
_____________________________________________________
АВСТРАЛОПИТЕК ГАРХИ
Считается, что австралопитек garhi
существовать между 2 и 3
миллионов лет назад в регионе среднего Аваша.
Эфиопии.Первый
частями гархи, которые должны были быть обнаружены, были череп, зубы и кости конечностей
и были найдены Берханом Асфау и Тимом Уайтом. Это был значительный
и удивительная находка, потому что у Гархи был маленький мозг, но он использовал и сделал
инструменты для взлома костей конечностей животных, чтобы добраться до
богатый питательными веществами костный мозг в центре кости. Соответственно, слово
гархи означает удивление на местном афарском языке Эфиопии.Перед
открытие A. garhi, первыми пользователями инструмента считались
Род Homo. А. Гархи, таким образом, считается связующим звеном
между
Australopithecus и Homo из-за маленького мозга / инструмента, использующего
комбинация. A. garhi, по-видимому, существовал во время гипотетического
туннель
времени, из которого виды появились примерно через 3 миллиона лет, не используя
инструменты, не очень хорошо адаптированные к окружающей среде и с маленьким мозгом
и появился около 2 миллионов лет назад с помощью инструментов, хорошо приспособленных к
их окружение, и с большим мозгом.
www.modernhumanorigins.com/garhi.html
http://www.ashtonfarm.demon.co.uk/early_hominid_evolution/a_garhi.htm
http: // biology.about.com/library/weekly/aa042999htm
Половой диморфизм у Australopithecus afarensis был подобен таковому у современного человека
Abstract
Значительная летопись окаменелостей для Australopithecus afarensis включает как частичный скелет взрослой особи [местонахождение в Афаре (A.Л.) 288-1, «Люси»] и большое собрание одновременных погибших (A.L. 333). Здесь мы оптимизируем данные, полученные от обоих, для более точной оценки размера скелета диморфизм. Посткраниальные соотношения, полученные из A.L. 288-1, позволяют значительно увеличение размера выборки по сравнению с предыдущими исследованиями. Обширное моделирование использование современных людей, шимпанзе и горилл подтверждает, что этот метод точным, и этот диморфизм размеров скелета у A. afarensis был наиболее вероятным. аналогичен современному Homo sapiens .Эти данные исключают некоторые очевидные несоответствия между диморфизмом размеров клыка и скелета у гоминоидов, подразумевают, что этот вид не характеризовался существенным половым биматурации и значительно увеличивают вероятность того, что репродуктивная Стратегия A. afarensis была в основном моногамией.
Правильный вывод степени полового диморфизма у ранних гоминидов — это имеет решающее значение для понимания их палеобиологии. С момента выздоровления и диагностики ранних видов гоминидов Australopithecus afarensis в 1970-х гг., оценки его диморфизма занимали видное место в интерпретации его филогения и поведение (1–5).В результате многочисленных несистематических оценок в настоящее время широко считали, что A. afarensis был значительно более диморфным, чем современный человек (6). Некоторые имеют вышли за рамки этой ортодоксии, чтобы утверждать, что вариации по половому признаку превышают что можно увидеть у любого живого гоминоида (7). Ни один из этих анализов, тем не менее, адекватно компенсирует влияние временных и географических вариация в противоположность нормативному диморфизму на уровне популяции. Так как анатомическая структура развивается со временем, обязательно, чтобы половые диморфизм внутри вида не следует отождествлять с вариациями, увеличенными за счет эволюционное изменение.Здесь мы используем современную Афарскую местность (A.L.) 333 образец гоминид, чтобы показать, что диморфизм в A. afarensis маловероятен превзошли у современных людей.
Реконструкция диморфизма размеров тела в окаменелостях подлежит ошибки сложения ( i ) малых выборок и ( ii ) потенциальное смешение экогеографических и / или временных вариаций с этим связаны с сексом. Несмотря на убедительные дискуссии в пользу исключительной осторожности в оценка раннего полового диморфизма гоминидов по этим причинам (8–10), наиболее часто цитируемые оценки по-прежнему основаны на простых соотношениях масса тела, предсказанная для нескольких изолированных особей, пол которых был оценен как a priori (т.е., исходя из размера). Эта круговая практика имеет методически исключенные промежуточные образцы (Рис. 1) и сильно ограничены размер образца. Действительно, наиболее часто цитируемая оценка (см., Например, исх. 11) диморфизма в A. afarensis (44,6 ± 18,5 для самцов; 29,3 ± 15,7 для женщин) полагались только на трех предполагаемых женщин, две из которых входят в наименьшие экземпляры, зарегистрированные для данного вида (6). Тем не менее, из-за небольшого размер выборки, 95% доверительный интервал все еще включает мономорфизм (3).
Рис.1.Образец некоторых оригинальных экземпляров дистального отдела плечевой кости из A. афарский. ( A ) A.L. 288-1; ( B ) A.L. 322-1; ( C ) А.Л. 137-48А; ( D ) A.L. 333-29; и ( E ) MAK VP 1/3. Просто Несистематическое сравнение A и E дает очень высокий диморфизм; однако наличие других экземпляров среднего размера проверяет, что такие оценки не могут быть правильными (см. Таблица 1).
Систематическая случайная выборка (например, «бутстрэппинг») скелетных размеры — это более мощная и более точная основа для оценки половой диморфизм в ископаемых образцах (12–17). Такие процедуры сравнивают предписанные индексы [например, максимальные / минимальные отношения (MMR) или коэффициенты вариации (CV)] для комплексов окаменелостей к тем созданы из образцов дошедших до нас видов. Хотя, безусловно, лучше, чем меньше систематические подходы, эти новые методы все еще страдают от проблемы, заключающейся в том, что скопления окаменелостей редко сохраняют достаточное количество примеров одних и тех же анатомических элемент.Кроме того, даже с большими выборками остается вероятность смешения экогеографических и временных вариаций размера тела с этим из-за секса. По умолчанию существующие образцы гоминоидов, которые обеспечивают стандарты, с которыми сравниваются коллекции окаменелостей, все современные, тогда как ранние окаменелости гоминидов обычно накапливались как минимум несколько сотен тысяч лет (например, см. 1 и 14 и ниже). Четный ненаправленные колебания размеров тела со временем увеличат размерный диапазон любого образца окаменелости (например,g., лица, изображенные на Рис.1), и как географические, так и географические. и временные факторы с особой вероятностью повлияют на оценки, сделанные с использованием MMR (12, 13, 16). Таким образом, мы почти всегда сталкивался с вопросом, можно ли оценить диморфизм ископаемых отражает истинный биологический диморфизм или вместо этого указывает на вариацию, усиленную комбинированная неоднородность географии, времени, экологии и даже видов сам состав.
Материалы и методы
Эти проблемы потенциально решаемы для A.afarensis , потому что его летопись окаменелостей включает большое скопление геологически одновременной смерти из одного слоя в одной местности (A.L. 333) (18). Следовательно, нет а априорные предположения относительно потенциального воздействия таких факторов являются требуется, и этот сайт представляет уникальную возможность изучить их в ранние виды гоминидов. Различные посткраниальные фрагменты, извлеченные при этом Встречи, однако, представляют собой множество различных анатомических элементов. К счастью, частичный скелет взрослого человека (A.Л. 288-1, «Люси») также доступны для этого вида. Поэтому мы использовали отношения между одним размер скелета [диаметр головки бедренной кости (FHD)] и другие показатели скелета сохранено в A.L. 288-1 для прогнозирования FHD для 22 экземпляров, извлеченных на A.L.333 (см. Оценку точности ниже) и семь дополнительных образцов из другие населенные пункты Хадар и Мака [Объединенный Афар (CA)] (Таблица 1). A.L. 288-1 также сохраняет нижнюю челюсть, но мы не использовали ее здесь по двум причинам: ( i ) только три поддающихся измерению взрослых особей были доступны от A.Л. 333 и ( ii ) посткрании являются лучшими коррелятами размера тела; действительно, точность оценки диморфизма размеров тела по данным нижней челюсти кажется сомнительным (см. ниже). FHD был выбран из-за его широкого использования в оценка размера. Наши результаты были бы идентичны, если бы мы стандартизировали любой другой размер скелета, потому что все соотношения и меры дисперсии [то есть, индекс биномиального диморфизма (BDI), CV и MMR] останется одно и тоже.
Таблица 1. А.Пример afarensis , используемый для моделированияМы рассчитали три меры скелетного диморфизма для A.L. 333 и C.A. образцы: MMR, CV и BDI; ранее техника диморфизма (19). Последний из них требует трех допущений: ( i ) оба пола присутствуют в выборке; и ( ii ) любой экземпляр имеет равную априорную вероятность быть самцом или самка, но ( iii ), когда два экземпляра разного пола, более крупный мужчина. Чтобы применить этот простой алгоритм, все образцы сначала собираются в зависимости от увеличения размера.Тогда имеется n — 1 возможный пол распределения (от одной женщины / все остальные мужчины к одному мужчине / всем остальным женщинам), и n — 1 отношений, для которых оценка полового диморфизма может быть рассчитано (среднее предполагаемых самцов / среднее предполагаемых самок). BDI — это средневзвешенное значение этих n — 1 оценок с использованием каждого отношения вероятность в биномиальном разложении. Остальные показатели (CV и MMR) были рассчитывается, как описано (12, 13). Мы использовали небольшую выборку поправка на CV (20).К судить о точности этих трех методов (BDI, CV и MMR), мы их применили к идентичным анатомическим массивам скелетных показателей, случайно выбранных из сохранившиеся образцы Homo, Pan и Gorilla (Таблица 2) (т.е. точно такая же метрики, рассчитываемые для A.L.333 и расширенного C.A. образец). Мы сделали так 1000 раз для каждого таксона. На каждой итерации образец шаблона (действующий в роль A.L. 288-1) был выбран случайным образом, чтобы обеспечить соотношения для оценки FHD. Поскольку размеры контрольных образцов гоминоидов конечны ( n ≈ 50, таблица 2), любой отдельный человек в выборке был выбран несколько раз в качестве шаблон.Однако каждый случайно выбранный анатомический массив ( n = 22 или 29, представляющих 12 различных показателей) почти наверняка уникальны для каждого из 1000 итераций.
Таблица 2. DSD фактических DM и соответствующие значения BDI, CV и MMR для сохранившиеся образцы гоминоидов, использованные в этом исследованииПомимо трех индексов полового диморфизма (см. Выше), мы также рассчитал два показателя фактического скелетного диморфизма (т.е. среднее мужское / женское среднее значение на основе известного пола) для каждого моделирования: первое с использованием FHD, оцененное как шаблонный метод для каждого образца [шаблонный половой диморфизм (TSD)], и второй с использованием фактических FHD для каждого человека [мы будем ссылаться на последний как прямой половой диморфизм (DSD)].Сравнение этих двух методов позволяет оценить потенциальную ошибку, возникающую при использовании шаблона образец, то есть сравнение TSD и DSD позволяет напрямую оценить предполагаемый (шаблонный) по сравнению с реальным (фактическим FHD) диморфизмом в каждом гоминоиде моделирование. Потому что расчет полового диморфизма для любого гоминида образцы (например, BDI, CV, MMR) требует использования образца-шаблона (A.L. 288-1), их можно оценить только путем сравнения с TSD из симуляции.
Для A.L. 333, мы столкнулись с тафономической проблемой, когда <22 взрослых могут быть представлены. При использовании зубных рядов нижней челюсти взрослого взрослый минимум количество человек в АЛ 333 было девять (16). Поэтому мы провели дополнительное моделирование, в котором количество гоминоидов, служащих источник 22 показателей в таблице 1 изменялось от 22 до 9. Как показано на Рис.2 A , поэтому ограничение количества взрослых существенно не повлияло на исход моделирования.
Фиг.2.( A ) Гистограммы, созданные путем моделирования выборки A.L.333 для 22 показатели обрабатываются отдельно и для 22 показателей, полученных случайным образом только из девяти индивидуумы (по 1000 итераций). Стрелка и вертикальная линия на каждом графике указывают на диморфизм для образца A. afarensis из A.L. 333. Примечание. что ограничивает источник этих показателей только девятью людьми ( минимальное количество человек в АЛ 333 — девять) не оказывает существенного влияния об оценке диморфизма.( B ) Гистограммы, созданные для образцов 22 (A.L. 333) и 29 (C.A.) метрики. Обратите внимание, что поведение BDI и CV почти идентичны, но MMR сильно переоценивают диморфизм (см. Таблица 3). Тем не менее, MMR предсказывают такую же степень диморфизма у A. afarensis , что значительно больше для C.A. образца, чем для A.L. 333, но все же большинство похожи на людей.
Результаты
Как и ожидалось, наше моделирование (см.рис.2 и Таблица 3) показывают диморфизм размеров тела у человека должен занимать промежуточное положение между недиморфными Пан и высокодиморфный Gorilla . По большей части шаблонный метод имеет тенденцию переоценивать как средние, так и дисперсии фактических значения диморфизма (сравните TSD с DSD в Таблица 3). В резюме нет дают прямую оценку диморфизма, но его поведение близко имитирует что BDI, что делает. MMR постоянно не позволяли существенно различать Пан от Gorilla и сильно переоцененный диморфизм во всем три таксона (рис.3 и Таблица 3). Их очень бедные производительность, вероятно, является результатом нестабильности самого метода, но была усиливается за счет использования шаблона (как отмечалось ранее, использование соотношений увеличивало разброс оценок). Однако даже если шаблон не использовался, а только были использованы фактические значения головки бедренной кости, распределение MMR по-прежнему переоценил диморфизм и не смог существенно различить Pan из Горилла . Эти методологические проблемы, возможно, не были очевидно до сих пор, потому что MMR обычно применялись только к очень небольшие образцы.В отличие от MMR, как BDI, так и CV имеют адекватную количественную оценку. диморфизм образца, о чем свидетельствуют корреляции как с TSD, так и с прямым половой диморфизм (см. вспомогательную информацию, опубликованную на Веб-сайт PNAS, www.pnas.org). Оба были практически идентичны во всех симуляциях.
Рис.3.Средние абсолютные различия (и стандартные отклонения) между моделями фактический диморфизм (DM) и BDI и MMR с использованием шаблонного метода (TSD) для C.A. моделирование (таблица 3).BDI и MMR — это два метода в нашем исследовании, которые производят прямые оценки диморфизм. BDI переоценивает диморфизм шимпанзе, потому что этот вид фактически недиморфный. Для видов с диморфизмом выше Homo и Gorilla , однако фактический DM находится в пределах диапазона 2-SD индексы диморфизма.
Таблица 3. Результаты моделированияBDI и CV рассчитаны для A. afarensis , будь то из A.L. 333 или весь C.A. образец, были наиболее совместимы с Homo симуляции. Что касается Pan , только C.A. BDI и CV были значительно больше, чем смоделированное распределение. Однако оба гоминида образцы значительно отличались от Gorilla (на самом деле A.L. 333 BDI полностью выпал за пределы диапазона смоделированных значений) (Таблица 4). Решающего Важным является тот факт, что C.A. образец дал значительно более высокий диморфизм (например, BDI = 1,222), чем A.L.333 (BDI = 1.167) (Рис.2 B ), подразумевая что C.A. выборка отражает не только половой диморфизм, но и экогеографические и временные факторы.
Таблица 4. Точный подсчет количества значений моделирования, которые упали меньше (<) или больше (>), чем значение A. afarensisОбсуждение
Эти результаты соответствуют недавнему исследованию размера нижней челюсти в A. afarensis на изменения с течением времени (14). Его авторы имели Ранее был сделан вывод, что А.afarensis мог быть таким же диморфным как Gorilla и Pongo (13). Однако их посткраниальные образцы были слишком малы (бедренная кость = 5 и плечевая кость = 3) для расчета CV, и вместо этого они использовали MMR. Поскольку MMR чрезвычайно чувствителен к географическому и временные вариации, только за счет включения данных от всех отобранных индивидуумов в расчет индекса (например, CV или BDI) можем более надежно оценить вариация, которая объясняется половым диморфизмом. Их большая выборка для тела нижней челюсти [ n = 17 (среднее геометрическое высоты нижней челюсти и толщина M 1 )] дала CV, равный 11.7 для А. afarensis [по сравнению с 12,3 для (очень диморфных) горилл и 7,78 для (недиморфные) шимпанзе]. Однако последующая амплификация этого образца (до n = 20) и удаление четырех самых последних образцов (все еще сохраняя временной диапазон ≈320000 лет) уменьшил А. afarensis CV до 8.49 (14), что значительно ниже, чем для сопоставимые показатели в изолированных современных популяциях людей, таких как Зулу (10.2) и Спиталфилдс (10.1) (21).Более того, как уже отмечалось ранее оценка диморфизма размеров тела на нижней челюсти могла быть необоснованно. Тот же показатель во второй независимой выборке (21) дали практически идентичные Резюме для людей (10,4) и горилл (10,5). Если бы использовалась только высота корпуса (что предположительно более прямо отражает диморфизм собак), нижние челюсти показал лучший результат (13,9 у гориллы против 11,6 у человека), но у шимпанзе женское среднее было больше, чем у мужчины. § В любом случай, если четыре последних А.afarensis особи снова исключено из расчетов (см. выше), CV высоты тела нижней челюсти (10.2) (14) все еще падает ниже что у современных людей.
Мы обратились к еще одной потенциальной тафономической проблеме. Наименьший расчетное значение FHD для A.L. 333 составило 29,8 (Таблица 1). Хотя это значение чуть более чем на 1 мм больше, чем «Люси» (28,6), это могло гипотетически означать недопредставленность самок размером «Люси» на участке (хотя Маленькие молодые особи систематически систематически искажают сохранение / восстановление у A.Л. 333 очень маловероятно). Тем не менее, мы последовательно увеличивали наш A.L. 333 образец с образцами размера «Люси» (то есть, FHDs 28,6), пока его BDI сравнялся с гориллами. Это потребовало добавления восьми «Люси» по сравнению с предыдущим общим числом 22 (т. Е. Новым n = 30 всего элементов). Однако добавление этих гипотетических элементов также увеличивает минимальное количество особей в 333 г. всего от 10 (первоначальные девять плюс один, представленный восемью новыми элементов) и 17 (исходные девять плюс восемь новых особей, каждый представлен одним элементом) и увеличивает выборку показателей более чем на треть.Очевидно, что это искусственное и нерепрезентативное раздувание реальный образец A.L.333. Более того, нет априори причин для предполагают, что крошечные экземпляры, такие как A.L. 288-1, представляют средний размер из самок A. afarensis . В самом деле, если они это сделали, большинство промежуточных тогда особи будут самцами, а самцы A. afarensis размером тела будут тогда будет гораздо более изменчивым, чем у любого другого живущего гуманоида. Аналогично любой утверждение, что A.L. 333 представляет собой одного полигинного мужчины, сопровождаемого несколько самцов и молодых особей потребуют уникальных экстремальных различий в размерах самки.Более того, если бы соотношение полов было таким же, как у большинства групп приматов, (варьируется от примерно равного соотношения полов до единственного мужской), диморфизм A.L.333 должен был быть в пределах или ниже современных человеческий диапазон (рис. 4).
Рис.4.Простое соотношение мужчин и женщин FHD (с использованием A.L. 288-1 в качестве шаблона) для A.L. 333 (для Комбинированного Хадара см. Вспомогательную информацию) в зависимости от количество сохранившихся самок [т.е. полный диапазон всех соотношений, используемых для рассчитать BDI (TSD в таблице 3)].Диморфизм уровня горилл присутствовал на этом участке только в том случае, если в нем была обнаружена единственная особь женского пола. Во всех остальных случаях (самка n = 2–21), диморфизм был близок или намного ниже среднего современного человеческого BDI. (оценка TSD; таблица 3). Этот расчет предполагает, что самцы всегда крупнее самок, т.е. почти наверняка неверен на этом уровне вероятного диморфизма и, следовательно, также является завышенной оценкой действительного диморфизма. Действительно, если более половины выборка — самка, ее BDI (рассчитанный TSD и, следовательно, завышенный) все еще очень близки к человеческим ценностям актуального (известного пола) диморфизма (см. Таблица 3).
Свидетельства того, что A. afarensis характеризуется лишь незначительной умеренная степень скелетного диморфизма разрешает парадокс, сформулированный Плавкан и ван Шайк (4), которые отметил, что в основном мономорфные клыки A. afarensis делают его предполагаемый большой диморфизм массы тела (снова опираясь на исх. 6) трудно интерпретировать (см. также исх. 24). Они также заметьте, что это не обязательно загадка, потому что диморфизм массы тела часто не отражает конкуренцию между мужчинами, но может возникать из-за других такие факторы, как предпочтения субстрата, избегание хищников и филогенетические инерция.Тем не менее, вопрос во многом решается по результатам Настоящее исследование и особенно по распределению размеров скелета представлены на рис. A.L.333 и весь образец A. afarensis (см. Также Рисунок 1). Эти распределения полностью несовместимы с выраженным биматуризмом, как у горилл и орангутангов [то есть телическая гетероморфоза Джармана (25)], которому А. afarensis постоянно сравнивали в прошлом. Вместо этого они сравнимо с гомеоморфным диморфизмом, который характерен для людей и шимпанзе.
Произведенные гоминиды обладают рядом уникальных / необычных персонажей, в том числе скрытая овуляция (у обоих полов) (26), сложные эпигамии в оба пола, относительно небольшие семенники (по сравнению с массой тела), относительно короткая сперма (26), навсегда увеличенные молочные железы и резко увеличенная церебральная способность, не имеющая аналогов в другие млекопитающие (27). Оно имеет Было высказано предположение, что все эти персонажи, кроме последнего, происходят из социальных комплекс, в том числе обеспечение мужчин, обусловленное выбором женщин, что позволило Australopithecus для противодействия ограничению скорости воспроизводства вызванный чрезмерным K-отбором (28).Хотя инструмент, использующий утверждалось, что сценарии объясняют уменьшение количества собак, например «Неиспользованные» модели не могут адекватно объяснить выбор направления для уменьшения коронки у мужчин и множества других уникальных анатомических знаков производных гоминидов. Одна очевидная возможность состоит в том, что самцы с маленькими клыками будут менее эффективными конкурентами в полигинной стратегии спаривания и будут тем самым оказываются более надежными поставщиками (и тем самым дифференцированно выбирают самки). Однако женский выбор не дает адекватного объяснение паттернов скелетного диморфизма, наблюдаемого у гоминоидов.В первую очередь потому, что у шимпанзе практически нет значительного скелета. размерный диморфизм (диморфизм массы тела умеренный), несмотря на их явно выраженные полигинная репродуктивная стратегия. Этот почти мономорфизм резко контрастирует к заметному диморфизму скелета горилл и орангутангов и предполагает, что скелетный диморфизм сам по себе является плохим предиктором репродуктивной стратегии в гоминоиды.
У полигинных видов, у которых самцы должны полагаться на значительную массу тела и оружие (особенно большие клыки) для борьбы за доступ к товарищам, это типично для задержки созревания самцов и, следовательно, продления роста чтобы подчеркнуть эти характеристики (телический гетероморфизм) (25, 29, 30).Потому что гоминоидный самец размер клыков во многом является результатом длительного роста (31), большее время, чтобы зрелость у горилл и орангутангов (29, 30, 32) соответствует большая индивидуализация успеха самцов у этих видов, что отражено в их более выраженный диморфизм клыков, скелета и массы тела.
Родственные мужские сообщества P. troglodytes сильно зависят от территориальное сопровождение «патрулями». Поэтому отбор может иметь способствовали более быстрому созреванию скелета самцов, чтобы ускорить их участие в кооперативной территориальной защите (это также будет учитывать видовые снижение диморфизма собак по сравнению с горилл и орангутангов) (33). Сковорода панискус есть слегка биматурационный из-за более позднего прекращения роста у самцов. Ли и Ши (30) атрибут эта большая биматурация снижает конкуренцию за кормление среди самок в P. paniscus , но не менее вероятным объяснением является то, что бонобо вместо этого представляют примитивное состояние, а самка P. troglodytes имеют немного задержку полового созревания (тем самым устраняя биматурацию в вида) из-за их более интенсивной самки-самки соревнование.
В любом случае минимальное проявление биматурации у обоих видов увеличивает вероятность того, что группы австралопитеков не были полигинны, если выставили женскую систему передачи. Как Хамада и Удоно (33) утверждали, что « социальная система и экология предков человека, которые развили характерную модель роста должна была отличаться от таковой у шимпанзе »(исх. 33, стр. 283). Во-первых, их заметный демографический успех и способность вторгаться в новые потенциально опасные местообитания убедительно свидетельствуют о том, что они, как и шимпанзе, обитали в смешанных группы (27, 28).Если бы такие группы тоже были (как шимпанзе), вовлеченные в значительную территориальную оборону, в основном благодаря родственные группы, столь же слабая степень скелетного диморфизма шимпанзе можно было бы ожидать, как и больший диморфизм собак.
Вместо этого умеренный скелетный диморфизм A. afarensis (больше чем Pan и менее Gorilla ) предполагает несколько более длинный период развития у мужчин по сравнению с женщинами и, следовательно, несовместимо с территориальной стратегией шимпанзе.В то же время это также заметно несовместима со стратегиями, подобными стратегиям горилл и орангутанов, у которых диморфизм скелета выражен гораздо сильнее. Следовательно, наличие умеренного скелетного диморфизма, такого как в современном человека и A. afarensis , а уменьшенный клык-самец полностью согласуется с парной репродуктивной стратегией у ранних гоминидов; что если их репродуктивная стратегия была подобна шимпанзе, гоминиды должны показать только минимальный скелетный диморфизм, или, если он был подобен орангутангу или горилле, они должны показать больший скелетный диморфизм.Диморфизм собак должен быть присутствует в любом случае. Скелетный диморфизм ранних гоминидов согласуется с еще один особый персонаж гоминида, отсутствие прорезывания у самцов у собак. задерживается и, таким образом, совпадает с соматическим созреванием (как и во всех других гоминоидные виды) (34, 35). Таким образом, наблюдаемые уровни диморфизм размеров тела у A. afarensis не означает, что моногамия менее вероятны, чем полигиния как фундаментальная социальная система этих ранние гоминиды.
Благодарности
Мы благодарим Gen Suwa, Richard J.Смит, Роберт Экхардт и Тим Уайт за подробные и критические обзоры этой рукописи. Луиза Хамфри, Крис Дин, и Крис Стрингер любезно предоставили неопубликованные данные, а Брайан Ричмонд и Сан-Хи Ли обратил наше внимание на ключевые ссылки. Кевин Керн внес к сбору данных и компьютерному программированию. Это исследование было поддержано Гранты Национального научного фонда SBR-9729060 и BCS-9919211 (на C.O.L.).
Сноски
↵ ‡ Кому должна быть адресована корреспонденция.Электронное письмо: olovejoy {at} aol.com.
Сокращения: А.Л., Афарская местность; BDI, индекс биномиального диморфизма; MMR, максимальное / минимальное соотношение; TSD — шаблонный половой диморфизм; CV, коэффициент вариация; FHD, диаметр головки бедренной кости; C.A., Комбинированный Афар.
См. Комментарий на стр. 9103.
↵§ Фактически, известный диапазон массы тела самок одного подвида шимпанзе ( Pan troglodytes troglodytes ) превосходит все известные самцы от другого ( Pan troglodytes schweinfurthii ) (22, 23).Если бы образцы от каждого были комбинированные (что могло произойти, например, при сборке ископаемых фрагментов из разных мест или периодов времени) диморфизм был бы переоценен.
- Поступила 5 января 2003 г.
- Copyright © 2003, Национальная Академия Наук
Australopithecus afarensis — Австралийский музей
Ключевые открытия окаменелостей
В 1970-х годах две группы по поиску окаменелостей начали обнаруживать свидетельства древних предков человека в Восточной Африке.Одна группа, возглавляемая Дональдом Йохансоном, работала в Хадаре в Эфиопии. Другая группа, возглавляемая Мэри Лики, находилась на расстоянии более 1500 километров в Лаэтоли в Танзании. Окаменелости, обнаруженные на этих двух участках, имели очень похожие черты и возраст, но они не соответствовали окаменелостям каких-либо видов, известных в то время. Поэтому в 1978 году для них было создано новое название вида — Australopithecus afarensis .
В настоящее время этот вид представлен несколькими сотнями окаменелостей из Восточной Африки.
Важные образцы
- «Люси» AL 288-1 — частичный скелет, обнаруженный в 1974 году Дональдом Йохансоном в Хадаре, Эфиопия. Этот относительно полный женский скелет, возраст которого составляет 3,2 миллиона лет, является самой известной особью этого вида. Ее прозвали «Люси» после песни «Люси в небе с бриллиантами» в исполнении The Beatles. Эта песня была очень популярна в то время, когда ее нашли. Люси была всего около 110 сантиметров в росте, но умерла уже взрослой.Она была двуногой, что означает, что она могла ходить на двух ногах, но, вероятно, она также проводила много времени, лазая по деревьям в поисках еды или убежища.
- Колено AL 129 1a + 1b обнаружено в 1973 году в Хадаре, Эфиопия. Когда было обнаружено это колено возрастом 3,4 миллиона лет, это была первая окаменелость, свидетельствующая о том, что наши предки ходили на двух ногах более трех миллионов лет. Открытие этой окаменелости побудило команду Дональда Йохансона вернуться в Хадар, где они в конечном итоге нашли «Люси» и сотни других окаменелостей этого вида.
- LH 4 - нижняя челюсть, обнаруженная в 1974 году группой Мэри Лики в Лаэтоли, Танзания. Эта окаменелость является типовым экземпляром или официальным представителем этого вида.
- «Первая семья» — группа из окаменелостей A. afarensis , обнаруженных в Хадаре, Эфиопия, в 1975 году. Она стала известна как «Первая семья», поскольку содержала окаменелости девяти взрослых и четырех детей. Какая-то неизвестная катастрофа охватила эту семейную группу, похоронив их всех одновременно. Эта важная находка позволила ученым получить представление о биологии и развитии одного ископаемого вида.Это также предоставило доказательства того, что этот вид жил небольшими группами на основе возможных семейных связей.
- «Селам» или «ребенок Люси» — частичный ювенильный скелет, обнаруженный в 2000 году в Дикике, Хадар, Эфиопия. Она жила 3,3 миллиона лет назад и умерла в возрасте 3 лет. Сохранились кости, которые никогда ранее не встречались у этого вида. Это самый ранний из когда-либо обнаруженных скелет молодых гомининов, и он должен предоставить фантастические возможности для того, чтобы узнать больше об этом виде и о том, как развивались наши ранние предки.
- «Старший брат Люси» по прозвищу Кадануумуу («большой человек» на Афаре) — частичный скелет мужчины, обнаруженный в Афаре, Эфиопия. Кость руки была впервые обнаружена в 2005 году, и в течение следующих четырех лет были восстановлены другие части, включая лопатку, ребра, шейный позвонок, таз, кости ног (целая большеберцовая и частично бедренная кость) и ключица. Этот человек был ростом около 1,6 метра (на 30% больше, чем «Люси») и жил около 3,6 миллиона лет назад.
Следы Лаэтоли
Эти окаменелые следы были обнаружены в Танзании, Восточной Африке и датируются 3 годом.6 миллионов лет назад. Ископаемые кости A. afarensis были найдены поблизости, поэтому предполагается, что они оставили следы.
Совершенно похожие на человеческие следы были оставлены гомининами, которые прошли через слой пепла, осевшего на земле после извержения далекого вулкана. Капли дождя сделали пепел влажным, и даже сейчас на некоторых частях слоя пепла все еще видны вмятины, оставленные этими каплями дождя. На первый взгляд кажется, что два человека шли бок о бок. Тот, что слева, был намного меньше другого и, возможно, был ребенком.При ближайшем рассмотрении мы видим, что отпечатки справа размыты и на самом деле были сделаны двумя взрослыми — один следовал за другим и наступал на отпечатки, оставленные первым. На полпути по тропе люди останавливаются и поворачивают налево, прежде чем продолжить. Солнце вскоре высушило влажный пепел, который затвердел, как цемент. Действующий вулкан продолжал выбрасывать пепел, пока слой толщиной до 20 сантиметров не покрыл и землю, и следы.
За миллионы лет образовались дополнительные отложения, а некоторые были размыты ветром и водой.В конце концов некоторые из шагов остались незамеченными. Затем удачливый охотник за окаменелостями наткнулся на них, и история начала раскрываться.
Следы имеют большое значение, поскольку они являются первым прямым доказательством (т.е. не ископаемыми костями) того, что наши предки 3,6 миллиона лет назад ходили прямо. Следы окаменелостей очень похожи на наши собственные. Они показывают, что пятка была первой частью стопы, которая упала на землю. Большой палец ноги был совмещен с другими пальцами и оставил глубокий отпечаток, показывающий, что каждый шаг заканчивался опусканием пальца вниз.Ступни также имели центральные арки, которые помогали телу совершать каждый шаг.
Распространение
Окаменелости были найдены в Хадаре в Эфиопии и Лаэтоли в Танзании, примерно в 1500 км от них.
Отношения с другими видами
Australopithecus afarensis обычно считается прямым предком человека. Он также считается прямым предком более поздних видов Australopithecus и всех видов рода Paranthropus .
Названия Praeanthropus africanus и Praeanthropus afarensis были предложены в качестве альтернативы исследователями, которые считают, что этот вид не принадлежит к роду Australopithecus .
Современники?
В 2015 году группа под руководством Йоханнеса Хайле-Селассие описала в журнале Nature новый вид A. deyiremeda (на афарском языке deyi означает «близкий», а средство правовой защиты — «родственник»).Возраст окаменелостей составляет 3,5–3,3 миллиона лет, и они были обнаружены в Ворансо-Милле в Эфиопии, недалеко от участков того же возраста, где было добыто особей A. afarensis . Если верно, A. afarensis был не единственным гоминином в Восточной Африке в то время.
Ископаемые останки, все обнаруженные в марте 2011 г., включают частичную кость верхней челюсти (голотип BRT-VP-3/1), две нижние челюсти (паратипы BRT-VP-3/14 и WYT-VP-2/10) и изолированный зуб P4 в фрагменте верхней челюсти (упомянутый препарат BRT-VP-3/37).Ключевые особенности включали передние скулы, трехкорневые премоляры и небольшие коронки на первых молярах. Были проведены сравнения с другими известными гомининами среднего плиоцена, такими как Kenyanthropus platyops и A. afarensis ; первооткрыватели полагали, что существует достаточно различий, чтобы оправдать определение нового вида. Другие не согласны, утверждая, что проводить сравнения с K. platyops проблематично (единственный череп был сильно искажен и, возможно, плохо реконструирован) или что небольшого размера выборки недостаточно, чтобы сделать такие важные выводы.Вместо этого они считают останки частью переменной популяции A. afarensis .
Независимо от того, представляют ли эти конкретные окаменелости новый вид или нет, становится вероятным, что A. afarensis был не единственным видом, обитавшим в то время в этой области. В 2012 году Хайле-Селассие объявил об открытии частичной стопы возрастом 3,4 миллиона лет (BRT-VP-2/73), обнаруженной в регионе Афар в Эфиопии. Очевидно, он не принадлежал к A. afarensis , но еще не был отнесен к виду.
Ключевые физические особенности
Окаменелости показывают, что этот вид был двуногим (мог ходить на двух ногах), но все же сохранил многие обезьяноподобные черты, включая приспособление к лазанию по деревьям, маленький мозг и длинную челюсть.
Размер и форма тела
- самки вырастали чуть более одного метра (105 — 110 сантиметров), а самцы были намного больше, примерно 150 сантиметров в высоту
- Грудная клетка имела конусовидную форму, как у обезьян
Мозг
- Мозг был маленьким, в среднем составлял около 430 кубических сантиметров и составлял около 1.3% от веса их тела
- реорганизация мозга могла начаться с некоторого увеличения частей коры головного мозга
Череп
- многие черты черепа были довольно похожи на обезьяньи, включая низкий покатый лоб, выступающее лицо и выступающие надбровные дуги над глазами.
- В отличие от большинства современных человекообразных обезьян, у этого вида не было глубоких бороздок за надбровными дугами, а спинной мозг выходил из центральной части основания черепа, а не из спины.
- самцов имели костяной гребень (сагиттальный гребень) на верхней части черепа для прикрепления огромных мускулов челюсти. У этого вида гребень был очень коротким и располагался ближе к задней части черепа.
- небольшая подъязычная кость (которая помогает закрепить язык и голосовой ящик), обнаруженная в молодом экземпляре, позволяет предположить, что A. afarensis имел голосовой аппарат, похожий на шимпанзе.
- полукруглый слуховой проход, похожий по форме на африканских обезьян и A. . africanus , предполагая, что этот вид не был таким быстрым и подвижным на двух ногах, как современные люди.
Челюсти и зубы
- челюсти и зубы были промежуточными между человеческими и обезьянами:
- челюсти были относительно длинными и узкими.В нижней челюсти зубы располагались рядами, которые сзади были немного шире, чем спереди. В верхней челюсти размещение последнего моляра приводит к тому, что ряды зубов изгибаются сзади.
- Зубы передних резцов были достаточно широкими.
- клыки были заостренными и длиннее остальных. Размер клыков был промежуточным между размером обезьян и людей. Как и у обезьян, у самцов клыки были намного больше, чем у самок.
- Между клыками и соседними зубами часто присутствовала щель (диастема).Эта обезьяноподобная особенность возникла между клыками и резцами верхней челюсти, а также между клыками и премолярами нижней челюсти.
- Премолярные зубы нижней челюсти имели бугорки обезьяноподобные (бугорки на жевательной поверхности). Передний премоляр, как правило, имел один большой бугорок (обезьяноподобный), а не два равных по размеру бугорка, как у людей.
- Задние коренные зубы были среднего размера и напоминали человеческие по форме «Y-5». То есть у них было пять бугорков, расположенных так, что бороздки между бугорками образовывали Y-образную форму.
Таз
- Таз был похож на человеческий, поскольку он был коротким и широким, но в нем отсутствовали детали, позволяющие людям ходить шагающей походкой
Конечности
- конечности имели человеческие черты, указывающие на способность ходить на двух ногах
- бедренные кости (бедренные кости), которые наклонены к колену
- колени с расширенными и усиленными наружными мыщелками
- арочные ступни и широкие пятки
- большие пальцы ног на одном уровне с другими пальцами и не противопоставлены
- обезьяна- похожие особенности, которые предполагают способность лазать по деревьям
- мощные руки с длинными предплечьями
- очень короткие бедренные кости
- длинные, изогнутые кости пальцев рук и ног.
- впадина лопатки, которая обращена вверх, как у обезьяны, а не в сторону, как у человека, но имеет другие сходства с человеческими лопатками
Образ жизни
Культура
Этот вид, вероятно, использовал простые инструменты, которые могли включать палки и другие недолговечные растительные материалы, обнаруженные в непосредственной близости. Камни могли также использоваться в качестве инструментов, но нет никаких доказательств того, что камни были каким-либо образом сформированы или изменены.Кажется вероятным, что они жили небольшими социальными группами, состоящими из мужчин и женщин, детей и взрослых. Самки были намного меньше самцов.
В 2010 году в Дикике в Эфиопии были обнаружены окаменелые кости с порезами, возраст которых составляет около 3,4 миллиона лет. Эти кости явно свидетельствуют о том, что каменные орудия использовались для удаления плоти и, возможно, для разбивания костей с целью получения костного мозга. Никаких реальных инструментов обнаружено не было, поэтому неизвестно, были ли «инструменты» преднамеренно модифицированы или это просто камни полезной формы.Хотя на этом месте не было обнаружено останков гомининов, исследователи полагают, что A. afarensis был ответственен за следы порезов, поскольку никаких других видов гомининов, относящихся к этому периоду, не было обнаружено в этом регионе.
Окружающая среда и диета
Этот вид обитал в различных средах обитания. Некоторые группы населения жили в саваннах или редколесьях, другие жили в более густых лесах у озер. Анализ их зубов, черепа и формы тела указывает на то, что диета состояла в основном из растений.Однако ископаемые кости животных с порезами, найденные в Дикике в 2010 году, были отнесены к этому виду, что позволяет предположить, что они, возможно, включали в свой рацион значительное количество мяса. Микроскопический анализ зубной эмали показывает, что они в основном ели фрукты и листья, а не семена и другой твердый растительный материал. Их конусообразная грудная клетка указывает на то, что у них были большие животы, приспособленные к относительно низкому качеству и высокому объему диеты. Положение сагиттального гребня к задней части черепа указывает на то, что передние зубы обрабатывали большую часть пищи.
Дополнительная литература:
Йоханнес Хайле-Селассие и др. (2015) «Новые виды из Эфиопии еще больше расширяют разнообразие среднего гоминина», Nature 521, 483-488
Йоханнес Хайле-Селассие и др. (2012) «Новый Нога гоминина из Эфиопии демонстрирует множественные приспособления к двуногому виду плиоцена », Nature 483, 565-569
Spoor, Fred (2015). «Палеоантропология: средний плиоцен переполнен». Природа 521, 432–433
преобразований тела и разума на JSTOR
Значительные изменения произошли в эволюции человека между 2 г.5 и 1,8 миллиона лет назад. Впервые появились каменные орудия труда, мозг расширился, тела увеличились, половой диморфизм в размерах тела уменьшился, пропорции конечностей изменились, щечные зубы уменьшились в размерах, а черепа стали делиться более уникальными чертами с более поздними Homo. Хотя два самых ранних вида Homo, H. habilis и H. rudolfensis, сохранили многие примитивные черты, общие с видами австралопитеков, они оба имеют общие ключевые уникальные особенности с более поздними видами Homo. Двумя наиболее заметными общими производными характеристиками были размеры мозга и жевательного аппарата по отношению к массе тела.Несмотря на общие производные признаки H. habilis и H. rudolfensis, одним неожиданным осложнением при переходе от австралопитека к Homo было то, что посткраниальная анатомия H. habilis сохранила многие характеристики австралопитека. Однако у H. rudolfensis, похоже, был более человеческий план тела, похожий на более поздние виды Homo. Следовательно, H. rudolfensis может быть связующим звеном между австралопитеками и Homo.
Информация о журналеЕжегодный обзор антропологии®, публикуемый с 1972 года, охватывает важные события в подобластях антропологии, включая археологию, биологическую антропологию, лингвистику и коммуникативные практики, региональные исследования и международную антропологию, а также социокультурную антропологию.Журнал важен для антропологов, этнологов, археологов, лингвистов и ученых в смежных областях.
Информация об издателеAnnual Reviews была основана в 1932 году как некоммерческое научное издательство, чтобы помочь ученые справляются с постоянно растущим объемом научных исследований. Всесторонний, авторитетные и критические обзоры ведущих ученых мира в настоящее время публикуются по 26 дисциплинам в области биологии, физики и социальные науки.Согласно рейтингу института «Импакт-фактор» для индекса научного цитирования научной информации каждый годовой обзор оценивается в верхней части соответствующей тематической категории или рядом с ней. Доступный для поиска заголовок и базу данных авторов и коллекцию рефератов можно найти по адресу https://www.annualreviews.org//. Веб-сайт также предоставляет информацию и цены на все печатные тома, онлайн-публикации, и переиздание сборников.
австралопитеков | Палеонтология вики | Фэндом
грацильных австралопитеков (представители рода Australopithecus ) (лат. Australis «юга», греч. Pithekos «обезьяна») — это группа вымерших гоминидов, которые тесно связаны с людьми.
Evolution
Грасиловые австралопитеки имели несколько общих черт с современными обезьянами и людьми и были широко распространены в Восточной и Южной Африке уже от 4 до 1,2 миллиона лет назад. Самые ранние свидетельства того, что гоминиды в основном двуногие, можно наблюдать на стоянке Лаэтоли в Танзании. Эти следы гоминидов удивительно похожи на следы современных людей, и их возраст положительно составляет 3,7 миллиона лет. До недавнего времени следы обычно классифицировались как австралопитеки, потому что это была единственная форма до-человека, которая, как известно, существовала в этом регионе в то время; однако некоторые ученые рассматривали возможность переназначить их еще не идентифицированным очень ранним видам рода Homo .
Australopithecus afarensis и Australopithecus africanus — одни из самых известных вымерших гоминидов. A. africanus раньше считался предком рода Homo (в частности, Homo erectus ). Однако были обнаружены окаменелости, отнесенные к роду Homo , которые старше A. africanus . Таким образом, род Homo либо отделился от рода Australopithecus в более раннее время (последний общий предок — A.afarensis или даже более ранняя форма, возможно Kenyanthropus platyops ), или оба независимо развились от еще, возможно, неизвестного общего предка.
Согласно проекту генома шимпанзе, линии человеческого ( Ardipithecus , Australopithecus и Homo ) и шимпанзе ( Pan troglodytes и Pan paniscus ) разошлись от общего предка примерно 5-6 миллионов лет назад. , если принять постоянную скорость эволюции.Однако недавно открытые гоминиды несколько старше, чем предполагают молекулярные часы. Sahelanthropus tchadensis , обычно называемый « Toumai », имеет возраст около 7 миллионов лет, а Orrorin tugenensis жил по крайней мере 6 миллионов лет назад; расположение сосцевидных отростков обоих указывает на то, что они были двуногими и поэтому отошли от общего предка намного дальше по эволюционному пути. Поскольку о них мало что известно, они по-прежнему вызывают споры среди ученых, поскольку молекулярные часы человека определили, что эволюционный раскол между людьми и шимпанзе произошел по крайней мере миллион лет спустя.Одна теория предполагает, что люди и шимпанзе однажды разошлись, а затем скрестились примерно через миллион лет после расхождения. [1]
По мере накопления молекулярных доказательств предположение о постоянной скорости оказалось ложным — или, по крайней мере, слишком общим. Однако, хотя молекулярные часы нельзя слепо предположить, что они верны, во многих случаях они все же верны, и их можно проверить. Например, пользователи молекулярных часов разрабатывают обходные решения, используя ряд статистических подходов, включая методы максимального правдоподобия и более позднее байесовское моделирование.
Морфология
Мозг большинства видов Australopithecus был примерно на 35% от размера современного человеческого мозга. Большинство видов Australopithecus были миниатюрными и изящными, обычно не выше 1,2 и 1,4 м (примерно от 4 до 4,5 футов) в высоту. В нескольких разновидностях австралопитеков наблюдается значительная степень полового диморфизма, что означает, что самцы крупнее самок. Современные гоминиды, по-видимому, не демонстрируют половой диморфизм в такой степени — в частности, современные люди демонстрируют низкую степень полового диморфизма, при этом самцы в среднем на 15% крупнее самок.У австралопитеков самцы могут быть на 50% крупнее самок. Новое исследование предполагает, что половой диморфизм может быть гораздо менее выраженным, чем этот, но по этому поводу все еще ведется много споров.
Видовые вариации
Хотя мнения расходятся относительно того, следует ли включать виды aethiopicus, boisei и robustus в род Australopithecus , в настоящее время в научном сообществе принято считать, что их следует отнести к отдельному роду, Paranthropus , который, как полагают, произошел от наследственной линии Australopithecus .Вплоть до последнего пятидесятилетия большая часть научного сообщества включала все виды, показанные справа, в один род. Paranthropus , будучи более массивным и крепким, также морфологически отличался от Australopithecus , и его специализированная физиология также подразумевает, что его поведение сильно отличалось от поведения его предка.
Эволюционная роль
Воссоздание A. afarensis из Лаэтоли (Американский музей) Шаблон: 3d alt
Летопись окаменелостей указывает на то, что Australopithecus является общим предком отдельной группы гомининов, которая теперь называется Paranthropus («крепкий australopithecines »), и, скорее всего, род Homo , который включает современных людей.Хотя интеллект этих ранних гомининов, вероятно, был не более сложным, чем у современных обезьян, двуногий рост является ключевым доказательством, которое отличает группу от предшествующих приматов, являющихся четвероногими. Морфология австралопитека опровергает то, что ученые считали ранее, а именно, что большой мозг предшествовал двуногию. Если A. afarensis был определенным гоминином, оставившим следы в Лаэтоли, это усиливает представление о том, что A. afarensis имел небольшой мозг, но был двуногим.Подобные ископаемые свидетельства ясно показали, что двуногие существа задолго до появления большого мозга. Тем не менее, остается спорным вопрос о том, как двуногие люди впервые возникли миллионы лет назад (некоторые концепции все еще изучаются). Преимущества двуногости позволяли освободить руки для захвата предметов (например, переносить еду и детенышей) и позволяли глазам смотреть поверх высокой травы в поисках возможных источников пищи или хищников. Однако многие антропологи утверждают, что этих преимуществ было недостаточно, чтобы вызвать двуногость.
Радикальные изменения в морфологии произошли до того, как появились грациозные австралопитеки; строение таза и ступни практически неотличимы от современного человека. Зубы ровные, как у современного человека с маленькими клыками; однако эволюция Paranthropus привела к появлению более толстых зубных рядов. Живя в саванне, австралопитеки столкнулись с одной конкретной проблемой. В то время они были самыми медлительными приматами, и многие из них стали жертвами хищных существ (львов и вымерших Dinofelis ).
Большинство видов Australopithecus были не более искусны в использовании инструментов, чем современные приматы, не являющиеся людьми, однако современные африканские обезьяны, шимпанзе и совсем недавно гориллы, как известно, использовали простые инструменты (например, раскалывали орехи камнями). и используя длинные палки, чтобы выкапывать термитов в курганах). Тем не менее, Australopithecus garhi , по-видимому, был наиболее продвинутым представителем этой линии с его предположительно более древними артефактами из каменных орудий, чем самый ранний член рода homo, известный до сих пор Homo habilis . Останки A. garhi были найдены вместе с инструментами и зарезанными останками животных, что свидетельствует о зарождении очень примитивной инструментальной промышленности. Это заставило многих ученых подозревать, что A. garhi может быть предком рода Homo . Однако дополнительные данные могут помочь антропологам и ученым определить истинных предков.
Диета
В предварительном исследовании окаменелых зубов австралопитека в 1979 году антрополог Алан Уокер предположил, что австралопитек мог быть фрукторианцем. [2] Однако новые методы изучения окаменелостей показали, что австралопитек , вероятно, был всеядным. В 1992 году изотопные исследования соотношения стронций / кальций в окаменелостях австралопитека и показали, что этот вид почти наверняка потреблял животных. Эти данные были подтверждены в 1994 году с помощью анализа стабильного изотропного углерода. [3]
Известные образцы
Список литературы
- Barraclough, G. (1989). Стоун, Н.(ред.): Атлас всемирной истории , 3-е издание, Times Books Limited. ISBN 0-7230-0304-1.
- Лики, Ричард (1994). Истоки человечества . ISBN 0-465-03135-8.
- Уайт, Тим Д., и др. . «Аса Исси, Арамис и происхождение австралопитека». Nature 440 (13 апреля 2006 г.), 883-89.