Методическая разработка интегрированного урока по русскому языку, литературе и информатике на тему «Устаревшая лексика в сказках А.С,Пушкина»
Муниципальное бюджетное общеобразовательное учреждение
средняя общеобразовательная школа № 20 пос. Зеленый
(МБОУ СОШ № 20 пос. Зеленый)
__________________________________________________________
142452 Московская область, Богородский городской округ, пос. Зеленый, д.4;
телефон: 8-916-130-29-31, e-mail: sosh30zelen@yandex.ru, официальный сайт: http://sosh30-zelen.edusite.ru/
Интегрированный урок
по русскому языку, литературе и информатике
по теме
«Устаревшая лексика в сказках А.С.Пушкина»
Работу представляет:
Симакова Екатерина Анатольевна,
учитель русского языка и литературы,
Дата рождения: 07.
Педагогический стаж работы: 3 года
Адрес электронной почты:
madam.mansurova@yandex.ru
Контактный телефон: 8(906)753-33-13
2019 г.
Аннотация
Методическая разработка урока по теме «Устаревшая лексика в сказках А.С.Пушкина» интегрирует знания из области русского языка, литературы и информатики, дает возможность применить полученные знания на практике.
На уроке используются наглядный, поисковый, аналитический методы. Формы организации познавательной деятельности: групповая, фронтальная. Урок должен проводиться в кабинете информатики, т.к. необходим 1 компьютер на группу из 3-4 человек.
Данные технологии обеспечивают не только интерес обучающихся, но и способствуют прочному закреплению знаний по предметам.
Разработка будет полезна преподавателям русского языка и литературы, а также информатики и ИКТ.
Работа состоит из следующих частей:
аннотации, в которой изложено краткое описание содержания и структуры разработки
основной части – технологической карты урока
приложения
Технологическая карта урока
Тема урока: «Устаревшая лексика в сказках А. С.Пушкина»
С.Пушкина»
Тип урока: комбинированный
Вид урока: урок-исследование
Методы обучения: наглядный, поисковый, аналитический
Формы организации: групповая, фронтальная
Цели урока:
обучающая – отработка умения определять типы устаревших слов, научиться пользоваться электронным словарем;
развивающая – развитие интереса к урокам русского языка, литературы, информатики, способности применять свои знания на практике;
воспитательная – воспитание любви к родному языку, формирование эстетического восприятия.
Задачи урока:
обобщить знания по теме «Устаревшая лексика»;
повторить и закрепить навыки работы в MicrosoftWord;
научиться использовать интернет-ресурсы для получения нужной информации.
Формируемые УУД:
личностные – формировать мотивацию учебно-познавательной деятельности, участвовать в творческом созидательном процессе;
регулятивные – определять тему и цели урока, составлять алгоритм действий в соответствии с поставленной задачей;
коммуникативные – уметь договариваться в группе, выстраивать связное высказывание.

Познавательные–находить и выделять необходимую информацию, осмысливать прочитанную или услышанную информацию.

Программно-дидактическое обеспечение: ЭВМ типа IBM, локальная сеть, ОС WindowsXP, MSoffise, проектор, тексты 7 сказок А.С.Пушкина
Предварительная подготовка: скопировать на каждый компьютер папки «Типы устаревших слов», распечатать кроссворды и оценочные листы.
Ход урока
Этапы урокаДеятельность учителя
Деятельность учащихся
Формирова-ние УУД
Организационный момент
Здравствуйте, ребята! Я рада вас видеть, улыбнитесь друг другу, я желаю вам успешной работы и хорошего настроения, садитесь.
Встают, приветствуют учителя, улыбаются друг другу, настраиваясь на позитивную работу.
Личностные –сформировать позитивный настрой;
Коммуникативные – умение слушать.
Актуализация знаний
Сегодня мы проведем с вами урок-исследование.
Молодцы! Расскажите, с помощью каких программ или приложений вы монтировали видео?
Сейчас мы проверим, насколько внимательно вы читали сказки А.С.Пушкина. У вас на столах лежат кроссворды (Приложение 2), посовещайтесь в группе и разгадайте его. Отлично, давайте проверим, обменяйтесь листочками.
Выходят группы учеников (3-4 человека), представляют короткие видео (буктрейлеры). Комментируют, чем именно понравилась сказка. Слушают друг друга. Рассказывают об использованных приложениях для монтажа – KineMaster, VivaVideo, VideoShow. Совещаются в группах. Разгадывают кроссворды по сказкам А.С.Пушкина. Проверяют, осуществляют взаимопроверку.
Личностные – осознание себя как личности и в тоже время как члена общества, коллектива;
Коммуникативные – учиться выражать свои мысли;
Познавательные – осмысление услышанной информации
Целеполагание
Ребята, посмотрите на экран. Перед вами предметы и слова, которые эти предметы изображают. Найдите соответствия. (Приложение 1). Откуда эти слова? Правильно, из сказок А.С.Пушкина. Как называются такие слова? Всегда ли мы знаем значение устаревших слов? Что могло бы нам помочь в определении лексических значений? Правильно словари. Как вы думаете, трудно ли создавать словари? Сегодня на уроке мы будем настоящими исследователями и продуктом нашего исследования станет созданный вами словарь устаревших слов из сказок А.С.Пушкина. Давайте сформулируем тему и цели урока.
Перед вами предметы и слова, которые эти предметы изображают. Найдите соответствия. (Приложение 1). Откуда эти слова? Правильно, из сказок А.С.Пушкина. Как называются такие слова? Всегда ли мы знаем значение устаревших слов? Что могло бы нам помочь в определении лексических значений? Правильно словари. Как вы думаете, трудно ли создавать словари? Сегодня на уроке мы будем настоящими исследователями и продуктом нашего исследования станет созданный вами словарь устаревших слов из сказок А.С.Пушкина. Давайте сформулируем тему и цели урока.
Изучают предметы и слова, представленные на экране проектора. Находят соответствия «предмет-слово». Определяют, что данные слова являются устаревшими и в современном русском языке не употребляются. Делают вывод, что для глубокого понимания содержания сказок Пушкина необходим специальный словарь, где будет собрана вся лексика из 7 сказок великого поэта. Определяют тему и цель урока-создание словаря.
Регулятивные – умение определять тему и цель урока
Обобщение и систематизация знаний
Ребята, посмотрите на рабочий стол вашего компьютера. Найдите папку устаревшие слова. Давайте вспомним и повторим, какие типы устаревших слов вам знакомы? Чем отличаются архаизмы от историзмов?
Найдите папку устаревшие слова. Давайте вспомним и повторим, какие типы устаревших слов вам знакомы? Чем отличаются архаизмы от историзмов?
Работают в группах за компьютером. Заходят в папку «Устаревшие слова», рассказывают все, что знают об устаревших словах, опираясь на схему.
Коммуникативные– умение общаться в группе, формулировать высказывание;
Познавательные – умение добывать информацию из схем.Составление алгоритма действий
Молодцы. Ребята, как вы считаете, с чего нам надо начать работу по составлению нашего словаря?
Составляют алгоритм действий: найти устаревшие слова в сказке, определить тип устаревшего слова, найти лексическое значение в словаре, расположить слова в алфавитном порядке.
Регулятивные – умение составлять алгоритм действий в соответствии с целью урока.
Физминутка
Сейчас вы будете работать с компьютерами, поэтому давайте выполним небольшую гимнастику для глаз.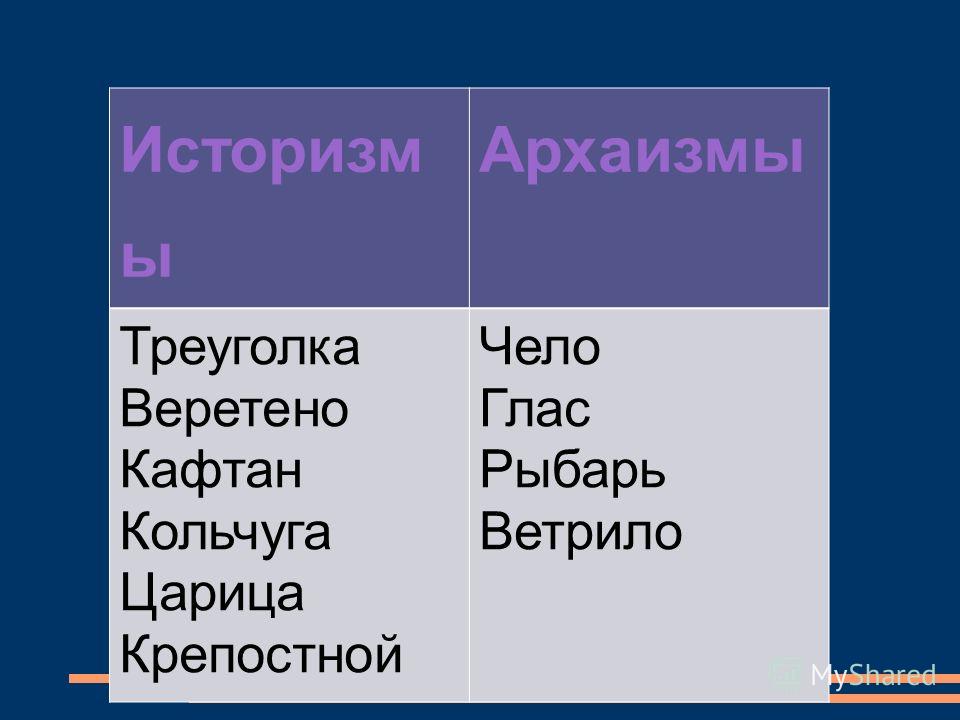
Выполняют гимнастику для глаз.
Работа в группах
Итак, решите, кто у вас в группе станет
аналитиком и будет выискивать устаревшие слова в сказке;
консультантом и будет определять историзм перед нами или архаизм;
Откройте браузер и в поисковой строке введите ссылку, которую видите на экране. Куда отправляет нас эта ссылка? Правильно, это электронный словарь устаревших слов (Приложение 5). Итак, приступайте к работе. Чья команда будет готова, поднимайте руки. Желаю удачи.
Распределяют роль в группе, работают по алгоритму в соответствии с поставленной задачей. Проходят по ссылке в электронный словарь устаревших слов, находят значение слов, определяют тип устаревшего слова, работают в программе Word.
Личностные – участие в творческом созидательном процессе;
Коммуникативные – умение договариваться в группе;
Познавательные – поиск и выделение новой информации, анализ объектов исследования.
Подведение итогов
Много слов удалось найти? Что нам осталось сделать? Работу по сбору и систематизации информации мы поручим
экспертам. Аналитики и консультанты дома поработают и представят на следующий урок полный словарь устаревших слов сказок А.С.Пушкина.Как вы думаете, кому полезен будет наш словарь? Как бы вы хотели, чтобы выглядело его оформление?
Остальные ребята дома выполнят другое задание – отгадают кроссворд по одной из предложенных тем «Архаизмы в сказках А.С.Пушкина». (Приложение 3 ) или «Историзмы в сказках А.С.Пушкина».
(Приложение 4)
Рассказывают о проделанной работе, приходят к пониманию того, что теперь осталось систематизировать и обобщить всю проделанную работу, чтобы получился настоящий словарь. Договариваются о внешнем виде словаря, решают прошить и подарить школьной библиотеке.
Коммуникативные –умение формулировать связное высказывание;
Регулятивные – умение прогнозировать предстоящую работу.
Рефлексия
Ребята, вы большие молодцы. Вы провели аналитическое исследование, поработали с электронными словарями, создали настоящий словарь. У вас на столах лежат оценочные листы (Приложение 6 ), заполните их, пожалуйста. Спасибо за урок. До свидания!
Заполняют оценочные листы.
Прощаются с учителем.
Регулятивные – умение определять степень успешности своей работы.
Приложение 1Нахождение соответствий «предмет-слово»: изразец, кичка, рогатка, калымага, чешуя, душегрейка, клёв, сарачинская шапка, печатный пряник, шелом
1
2
3
4
5
6
7
8
9
10
11
Приложение 2
Кроссворд по сказкам А. С.Пушкина
С.Пушкина
Вопросы по горизонтали:
1. Кто откусил кусочек отравленного яблока?
2. С каким предметом осталась старуха у ветхой землянки?
3. Кто охранял старого царя Дадона от нападения врагов?
4. Кто так говорил: «Дурачина ты, простофиля!»
5. Что не заплатили бесы попу?
6. Как звали царя, который женился на одной из трёх сестер?
7. Тот, кого выудил из моря Балда?
1. Что «под косой блестит» у царевны Лебедь?
2. Кто зашёл к царице с сыном в палату, огласить указ царя?
3. Кто помог царевичу Елисею найти царевну?
4. Кличка собаки из «Сказки о мёртвой царевне»
5. Какой титул был у Гвидона?
6. Кто помогал старику выполнять все желания старухи?
7. Сколько раз закидывал старик невод?
Приложение 3
Кроссворд
«Архаизмы в сказках А.С. Пушкина»
12
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
По горизонтали: 3-откуда;5-пальцы; 6- теплая короткая кофта без рукавов со сборками сзади; 9-но; 11- или; 12-плетеная веревочка, шнурок; 14-недомолвка; 15-воскликнуть; 16-вкусное кушанье; 17-восьмой; 19-откуда; 20-даже; 23-намеченная цель; 24-толкать в спину и бить по шее; 25-клюв; 26- золото;28- город; 29-разница.
По вертикали: 1-девушка; 2-мордочка; 4-встать на задние лапы; 7-сны; 8-но; 10- нежно заботиться; 13- позже; 17-глаза; 18- распространяющиеся слухи; 21-до сих пор; 22-звать; 27-злое дело.
Ответы
По горизонтали: 3-отколе;5-персты; 6-душегрейка; 9-ан; 11- аль; 12-снурок; 14-обиняк; 15-возопить; 16-яство; 17-осьмой; 19-отколь; 20-инда; 23-мета; 24-взамен; 25-клёв; 26-злато;28- град; 29-розь.
По вертикали:1-молодица; 2-мордка; 4-дыбы; 7-грезы; 8-ан; 10-холить; 13- ужо; 17-очки; 18- молва; 21-доселе; 22-кликать; 27-лихо.
Приложение 4
Кроссворд
«Историзмы в сказках А.С. Пушкина»
12
3
2
4
5
6
13
3
5
7
8
9
10
18
7
11
12
13
9
21
14
15
16
16
17
18
По горизонтали: 3-плитка, внешне похожая на кафель;6-участник праздничного театрализованного обряда; 7- жена казначея; 9-мера длины, равная 1 км 100 м; 11- карета; 14-земледелец, крестьянин; 15-владение царя; 16-богато украшенный женский парадный головной убор, наподобие шапочки; 17-высокий, с башенкой наверху, дом; 18- светлая девичья комната.
По вертикали: 1-храбрый воин, богатырь; 2-поле сражения; 4-деревяный ошейник с 4 острыми концами (рогами), мешал наказанному спать; 5- продавец вина в кабаке или в питейном доме; 8-мера длины, равная 71 см; 9- военачальник; 10- войско; 11- разукрашенная карета; 12- приближенные царю дворяне; 13-жена мелкого канцелярского чиновника.
Ответы
По горизонтали: 3-изразец; 6-скоморох; 7- казначеиха; 9-верста; 11- колесница; 14-смерд; 15-удел; 16-кичка; 17-терем; 18-светлица.
По вертикали: 1-витязь; 2-побоище; 4-рогатка; 5- целовальник; 8-аршин; 9- воевода; 10- рать; 11- колымага; 12- царедворец; 13-подъячиха.
Приложение 5
Устаревшая лексика в сказках А.С. Пушкина
Устаревшая лексикаСовременное значение
Тип устаревшей лексики
Аль
Или
Архаизм
Ан
Но
Архаизм
Арши́н
Мера длины, равная 71 см
Историзм
Ба́ять
Говорить
Архаизм
Белена́
Двулетнее растение с дурманящим запахом
Архаизм
Благода́рствуй
Благодарю, спасибо
Архаизм
Боя́ре, боярин
Богатые и знатные господа, приближённые царя
Историзм
Боя́рыня
Госпожа
Историзм
Брег
Берег
Архаизм
Була́т
Сталь особой обработки для изготовления оружия
Архаизм
Быль
То, что реально произошло
Архаизм
Вари́ть мёд
Изготовить хмельной напиток
Архаизм
Вене́ц
Царская корона
Историзм
Верста́
Мера длины, равная 1км 100м
Историзм
Весть
Вести́
Архаизм
Взаше́и
Толкать в спину и бить по шее
Архаизм
Вздо́рный
Ничтожный, глупый, неверный
Архаизм
Взыгра́лоретиво́
Забилось сердце
Архаизм
Взыска́ться
Обеспокоиться
Архаизм
Ви́тязь
Храбрый воин, богатырь
Историзм
Воево́да
Военачальник
Историзм
Возопи́ть
Воскликнуть
Архаизм
Во́пит
Причитает с плачем
Архаизм
Выдь
Выйди
Архаизм
Вы́молвить
Сказать
Архаизм
Вы́травить
Выгнать
Архаизм
Гоне́ц
Курьер, посыльный
Историзм
Го́рница
Комната
Историзм
Го́сти
Купцы, чаще всего, иноземные
Историзм
Град
Город
Архаизм
Грёзы ночи
Сны
Архаизм
Гумно́
Площадка в селе, где молотили зерно
Историзм
Дале
Дальше
Архаизм
Двор
Придворные, приближённые царя
Историзм
Дворя́не,дворян
Высшее сословие в государстве
Историзм
Досе́ле
До сих пор
Архаизм
Душегрейка1
Тёплая короткая кофта со сборками сзади
Архаизм
Дыбы́
Встать на задние лапы
Архаизм
Дьяк прика́зный
Служащий в государственном учреждении (приказе)
Историзм
Жезл
Символ царской власти
Историзм
Заве́сть
Завести
Архаизм
Зави́дела
Увидела
Архаизм
Заздра́вный ковш
Ковш с вином, который пускают по кругу под тост о здравии и счастье молодожёнов
Историзм
Зазо́рный
Постыдный
Архаизм
Заку́сливые
Острые
Архаизм
Зла́то
Золото
Архаизм
Знать
Значит
Архаизм
Зыбь
Волнение на море
Архаизм
Игу́мен
Настоятель мужского православного монастыря
Историзм
Извести́
Погубить
Архаизм
Изразе́ц
Плитка, внешне похожая на кафель
Историзм
И́нда
Даже
Архаизм
Казна́
Деньги
Историзм
Казначе́иха
Жена казначея
Историзм
Камча́
Узорчатая ткань
Историзм
Ки́чка
Богато украшенный женский парадный головной убор наподобие шапочки
Историзм
Клёв
Клюв
Архаизм
Кли́кать
Звать
Архаизм
Княги́нечка
Один из титулов в иерархии дворянства
Историзм
Князь
Один из титулов в иерархии дворянства
Историзм
Колесни́ца
Карета
Историзм
Колыма́га
Разукрашенная карета
Историзм
Кольцо́
На входной двери вместо ручки; им же стучали в дверь
Историзм
Коря́читься
Судорожно дёргаться
Архаизм
Кручи́ниться
Расстраиваться, горевать
Архаизм
Круши́ться
Расстраиваться
Архаизм
Купе́ц
Торговый человек
Архаизм
Курга́н
Могильный холм
Архаизм
Ли́хо
Злое дело
Архаизм
Ме́та
Намеченная цель
Архаизм
Молва́
Распространяющиеся слухи
Архаизм
Мо́лвить
Сказать
Архаизм
Молоди́ца
Девушка
Архаизм
Мо́рдка
Мордочка
Архаизм
Мудрёный
Непонятный, заумный
Архаизм
Мурава́
Яркая, зелёная трава
Архаизм
Наси́лу
С трудом
Архаизм
Науда́чу
На авось
Архаизм
Не прива́льный остров
Остров, возле которого не причаливали корабли
Архаизм
Не́быль
Придумка
Архаизм
Неве́дома
Неизвестная
Архаизм
Неве́стакра́сная
Красивая невеста
Архаизм
Невмо́чь
Не по силам
Архаизм
Не́где
Где-то
Архаизм
Недои́мка
Неуплаченный в срок оброк
Архаизм
Неука́занный товар
Запрещённый товар
Архаизм
Обиня́к
Недомолвка
Архаизм
Обмере́ть
Упасть в обморок
Архаизм
Обро́к
Дань, деньги
Историзм
Огру́зить
Отяжелить
Архаизм
Осержа́лася
Рассердилась
Архаизм
Осьмо́й
Восьмой
Архаизм
Отко́ль
Откуда
Архаизм
Отряса́ть
Избавиться
Архаизм
Отсе́ль
Отсюда
Архаизм
О́чи
Глаза
Архаизм
Пала́ты
Большие залы во дворце, или обширные богатые здания
Историзм
Парчо́вая кровать
Покрывало на кровати и полог из драгоценной ткани-парчи
Историзм
Пеня́ть
Укорять
Архаизм
Пере́чить
Возражать
Архаизм
Персты́
Пальцы
Архаизм
Побо́ище
Поле сражения
Историзм
Пова́длив
Способен
Архаизм
Подво́рье
Двор с хозяйственными постройками
Архаизм
Подья́чиха
Жена мелкого канцелярского чиновника
Историзм
Пожди́
Подожди
Архаизм
Покла́сть
Положить
Архаизм
Пола́ти
Настил из досок над печью
Историзм
По́лба
Особый сорт пшеницы
Архаизм
Полверсты́
Мера длины, равная 550 м
Историзм
Поразду́май
Подумай
Архаизм
Пред
Перед
Архаизм
Прекосло́вить
Возражать
Архаизм
Престо́л
Трон, особое кресло на возвышении, на котором сидел царь в торжественных случаях
Историзм
Прикуси́ла
Надкусила, откусила
Архаизм
Проме́ж
Между
Архаизм
Пря́никпеча́тный
Пряник с оттиснутым (отпечатанным) рисунком или буквами
Архаизм
Ра́тные дела́
Военные дела
Историзм
Рать
Войско, сражение
Историзм
Рехну́ться
Сойти с ума
Архаизм
Рога́тина
Палка с двумя острыми концами, обожжённая для прочности на огне
Историзм
Рога́тка
Деревянный ошейник с четырьмя длинными концами (рогами), мешал наказанному спать
Историзм
Розь
Разница
Архаизм
Сарачинская шапка
Чалма
Архаизм
Сарочи́н
Сарацин, кочевое племя арабских разбойников
Архаизм
Светёлка
Светлая комната, отделённая от кухни сенями
Архаизм
Светли́ца
Светлая девичья комната
Историзм
Се́ни
Холодный коридор между крыльцом и жилыми комнатами
Историзм
Се́нная девушка
Служанка для чёрной работы
Историзм
Скоморо́х
Участник праздничного театрализованного обряда
Историзм
Смерд
Земледелец, крестьянин
Историзм
Снуро́к
Плетёная верёвочка, шнурок
Архаизм
Спе́шить
Сбить противника с коня
Архаизм
Спи́ца
Шпиль на крыше
Историзм
Спозна́ть
Понять, узнать
Архаизм
Спуд
Положить под спуд — запереть
Архаизм
Сребро́
Серебро
Архаизм
Стан
Военный лагерь
Историзм
Ста́рче
Старик
Архаизм
Столбова́я
дворя́нка
Дворянка старинного и знатного рода
Историзм
(на) Сторо́же
На дежурстве
Архаизм
Ступа́й
Иди
Архаизм
Суда́рушка
Жена
Архаизм
Супоста́т
Противник, враг
Архаизм
Те́рем
Высокий, с башенкой наверху, дом
Историзм
Тихомо́лком
Незаметно
Архаизм
То́ком слёзы точи́ть
Слёзы льются ручьём
Архаизм
Толоко́нный лоб
Глупый человек, дурак
Архаизм
Торго́вый гость
Купец, чаще всего иноземный
Историзм
Трезво́нить
Распространять слухи
Архаизм
Тро́йка
Упряжка в три коня
Историзм
Тужи́ть
Горевать
Архаизм
Тяга́ться
Соревноваться, соперничать
Архаизм
Удел
Владение царя
Историзм
Ужо́
Позже
Архаизм
Учини́ть
Сделать
Архаизм
Хозя́ям
Хозяевам
Архаизм
Хо́лить
Нежно заботиться
Архаизм
Хорони́ться
Прятаться
Архаизм
Царедво́рец
Приближённые к царю дворяне, его ближайшее окружение
Историзм
Целова́льник
Продавец вина в кабаке или питейном доме
Историзм
Чай
Ну, надеюсь
Архаизм
Череда́
Очередь
Архаизм
Черни́ца
Монахиня, странница в чёрной одежде
Архаизм
Честны́е го́сти
Заслуживающие уважение гости
Архаизм
Чета́ми
Парами, попарно
Архаизм
Чешуя́
Кольчуга
Историзм
Чуде́сить
Совершать необдуманные поступки
Архаизм
Чупру́н
Чуб, хохол
Архаизм
Шамаха́нская царица
Кабардинская царица
Историзм
Шело́м
Шлем
Архаизм
Я́ство
Вкусное кушанье
Архаизм
Приложение 6
Оценочный лист
Собором это историзм или архаизм.

Предлагаемое слово «собором» является существительным, употребленным в творительном падеже, таким образом разберем начальное слово «собор». «Собор», согласно толковому словарю Ожегова имеет два значения: это и здание главной церкви в населенном пункте, и собрание жителей или служителей церкви. В любом случае, само слово имеет исторические корни и давно появилось в русском языке.
Архаизм или историзм
Архаизмом (в переводе с древнегреческого «устаревший») называют выражение или слово, которое вышло из употребления и в текущий момент находится в пассивном словаре. Однако, предмет, который обозначало данное слово существует и по сей день. Например: синематограф – кино, аэроплан – самолет и т.д. Большинство архаизмов в русском языке составляет старославянские слова – перст – палец, льзя – можно и т.д.
Историзмом называют явление или событие, ушедшее из повседневной жизни и более не употребляемое. Например, нэпман – предприниматель времен новой экономической политики 1920-30-х годов, боярин – представитель старославянской знати. Стоит отметить, что историзмы это прежде всего понятия, а не слова.
Стоит отметить, что историзмы это прежде всего понятия, а не слова.
Отличия между историзмами и архаизмами
Как лексические единицы, историзмы и архаизмы являются трудноразличимыми и необходимо осторожно подходить к их употреблению.
Главные отличия состоят в следующем:
- В художественной сфере историзм отражает правдивость определенной эпохи, а архаизм только подражает старинному стилю.
- Историзм – исчезнувшее явление, а архаизм – просто устаревшее название существующего предмета.
- Архаизмы входят в пассивный (малоиспользуемый) словарь человека, а историзмы могут находится в активном.
Выводы
Таким образом слово «собор», употребленное по отношению к храму или церкви не является ни архаизмом, ни историзмом, а просто имеет старославянские корни. До сих пор в официальном названии многих исторических строений и зданий культа оно употребляется, например «Собор Василия Блаженного». «Собор» же, употребленный для обозначения какого-либо собрания людей, является архаизмом.
42 — Российское Общество интеллектуальной истории
 Н. Гирин. Авангар
Н. Гирин. АвангарЮ. Н. Гирин. Авангар
Ю. Н. Гирин. Авангар
Ю. Н. Гирин. Авангар
Ю. Н. Гирин. Авангар
Ю. Н. Гирин. Авангар
Ю. Н. Гирин. Авангар
 yumpu.com/xx/document/view/52339780/42-/65″ title=»А. А. Турыгин. Билеф»>А. А. Турыгин. Билеф
yumpu.com/xx/document/view/52339780/42-/65″ title=»А. А. Турыгин. Билеф»>А. А. Турыгин. Билеф
А. А. Турыгин. Билеф
А. А. Турыгин. Билеф
А. А. Турыгин. Билеф
А. А. Турыгин. Билеф
А. А. Турыгин. Билеф
Л. В. Нургалеева. Эм
 yumpu.com/xx/document/view/52339780/42-/79″ title=»Л. В. Нургалеева. Эм»>Л. В. Нургалеева. Эм
yumpu.com/xx/document/view/52339780/42-/79″ title=»Л. В. Нургалеева. Эм»>Л. В. Нургалеева. Эм
Л. В. Нургалеева. Эм
Л. В. Нургалеева. Эм
Л. В. Нургалеева. Эм
О. Л. АКОПЯНМАКСИМ Г
О. Л. Акопян. Максим
О. Л. Акопян. Максим
 yumpu.com/xx/document/view/52339780/42-/93″ title=»О. Л. Акопян. Максим «>О. Л. Акопян. Максим
yumpu.com/xx/document/view/52339780/42-/93″ title=»О. Л. Акопян. Максим «>О. Л. Акопян. Максим
О. Л. Акопян. Максим
О. Л. Акопян. Максим
О. Л. Акопян. Максим
В. Н. Захарова. Зару
В. Н. Захарова. Зару
В. Н. Захарова. Зару
 yumpu.com/xx/document/view/52339780/42-/107″ title=»В. Н. Захарова. Зару»>В. Н. Захарова. Зару
yumpu.com/xx/document/view/52339780/42-/107″ title=»В. Н. Захарова. Зару»>В. Н. Захарова. Зару
В. Н. Захарова. Зару
Г. В. БАКУСФИЗИОЛОГ
Г. В. Бакус. Физиоло
Г. В. Бакус. Физиоло
Г. В. Бакус. Физиоло
Г. В. Бакус. Физиоло
 yumpu.com/xx/document/view/52339780/42-/121″ title=»Г. В. Бакус. Физиоло»>Г. В. Бакус. Физиоло
yumpu.com/xx/document/view/52339780/42-/121″ title=»Г. В. Бакус. Физиоло»>Г. В. Бакус. Физиоло
Г. В. Бакус. Физиоло
И. Е. Андронов. “Маг
И. Е. Андронов. “Маг
И. Е. Андронов. “Маг
И. Е. Андронов. “Маг
И. Е. Андронов. “Маг
 yumpu.com/xx/document/view/52339780/42-/135″ title=»И. Е. Андронов. “Маг»>И. Е. Андронов. “Маг
yumpu.com/xx/document/view/52339780/42-/135″ title=»И. Е. Андронов. “Маг»>И. Е. Андронов. “Маг
И. Е. Андронов. “Маг
И. Е. Андронов. “Маг
С. В. Федотов. Книжн
С. В. Федотов. Книжн
С. В. Федотов. Книжн
С. В. Федотов. Книжн
 yumpu.com/xx/document/view/52339780/42-/149″ title=»С. В. Федотов. Книжн»>С. В. Федотов. Книжн
yumpu.com/xx/document/view/52339780/42-/149″ title=»С. В. Федотов. Книжн»>С. В. Федотов. Книжн
С. В. Федотов. Книжн
С. В. Федотов. Книжн
И. Е. Рудковская. Ма
И. Е. Рудковская. Ма
И. Е. Рудковская. Ма
И. Е. Рудковская. Ма
 yumpu.com/xx/document/view/52339780/42-/163″ title=»И. Е. Рудковская. Ма»>И. Е. Рудковская. Ма
yumpu.com/xx/document/view/52339780/42-/163″ title=»И. Е. Рудковская. Ма»>И. Е. Рудковская. Ма
И. Е. Рудковская. Ма
И. Е. Рудковская. Ма
И. Е. Рудковская. Ма
И. Е. Рудковская. Ма
И. Е. Рудковская. Ма
И. Е. Рудковская. Ма
 yumpu.com/xx/document/view/52339780/42-/177″ title=»С. Р. Матвеев. Идея «>С. Р. Матвеев. Идея
yumpu.com/xx/document/view/52339780/42-/177″ title=»С. Р. Матвеев. Идея «>С. Р. Матвеев. Идея
С. Р. Матвеев. Идея
С. Р. Матвеев. Идея
С. Р. Матвеев. Идея
С. Р. Матвеев. Идея
С. Р. Матвеев. Идея
С. Р. Матвеев. Идея
 yumpu.com/xx/document/view/52339780/42-/191″ title=»О. В. Заиченко. Осно»>О. В. Заиченко. Осно
yumpu.com/xx/document/view/52339780/42-/191″ title=»О. В. Заиченко. Осно»>О. В. Заиченко. Осно
О. В. Заиченко. Осно
О. В. Заиченко. Осно
О. В. Заиченко. Осно
О. В. Заиченко. Осно
О. В. Заиченко. Осно
О. В. Заиченко. Осно
 yumpu.com/xx/document/view/52339780/42-/205″ title=»О. В. Заиченко. Осно»>О. В. Заиченко. Осно
yumpu.com/xx/document/view/52339780/42-/205″ title=»О. В. Заиченко. Осно»>О. В. Заиченко. Осно
О. В. Заиченко. Осно
О. В. Заиченко. Осно
О. В. Заиченко. Осно
О. В. Заиченко. Осно
О. В. Заиченко. Осно
О. В. Заиченко. Осно
 yumpu.com/xx/document/view/52339780/42-/219″ title=»Л. П. ЛАПТЕВАВ. И. ГЕ»>Л. П. ЛАПТЕВАВ. И. ГЕ
yumpu.com/xx/document/view/52339780/42-/219″ title=»Л. П. ЛАПТЕВАВ. И. ГЕ»>Л. П. ЛАПТЕВАВ. И. ГЕ
Л. П. Лаптева. В.И. Ге
Л. П. Лаптева. В.И. Ге
Л. П. Лаптева. В.И. Ге
Л. П. Лаптева. В.И. Ге
Л. П. Лаптева. В.И. Ге
Л.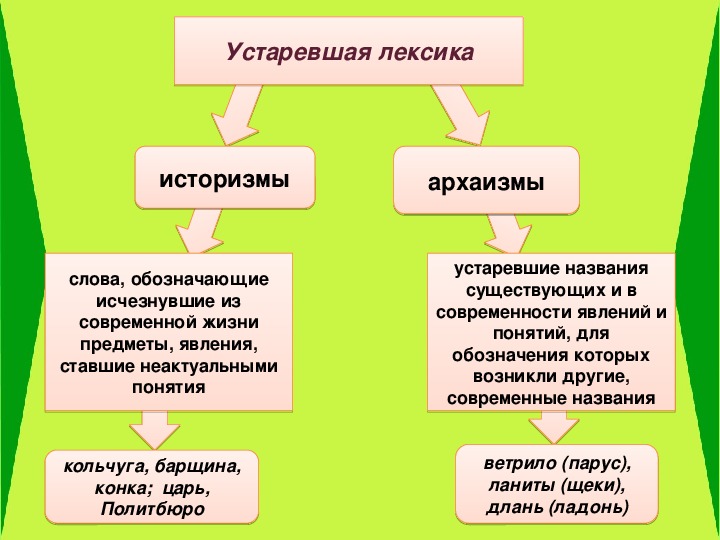 П. Лаптева. В.И. Ге
П. Лаптева. В.И. Ге
ИСТОРИЯ И ПАМЯТЬА.
А. В. Степанов. “На
А. В. Степанов. “На
А. В. Степанов. “На
А. В. Степанов. “На
А. В. Степанов. “На
 В. Степанов. “На «>А. В. Степанов. “На
В. Степанов. “На «>А. В. Степанов. “На
А. В. Степанов. “На
А. В. Степанов. “На
А. В. Степанов. “На
Н. А. Уткина. К истор
Н. А. Уткина. К истор
Н. А. Уткина. К истор
 yumpu.com/xx/document/view/52339780/42-/259″ title=»Н. А. Уткина. К истор»>Н. А. Уткина. К истор
yumpu.com/xx/document/view/52339780/42-/259″ title=»Н. А. Уткина. К истор»>Н. А. Уткина. К истор
Н. А. Уткина. К истор
Н. А. Уткина. К истор
Н. А. Уткина. К истор
В ПРОСТРАНСТВЕ ГЕН
В. С. Трофимова. Фил
В. С. Трофимова. Фил
 yumpu.com/xx/document/view/52339780/42-/273″ title=»В. С. Трофимова. Фил»>В. С. Трофимова. Фил
yumpu.com/xx/document/view/52339780/42-/273″ title=»В. С. Трофимова. Фил»>В. С. Трофимова. Фил
В. С. Трофимова. Фил
И. М. ЭРЛИХСОН“ТРЕТ
И. М. Эрлихсон. “Тре
И. М. Эрлихсон. “Тре
И. М. Эрлихсон. “Тре
И. М. Эрлихсон. “Тре
 yumpu.com/xx/document/view/52339780/42-/287″ title=»И. М. Эрлихсон. “Тре»>И. М. Эрлихсон. “Тре
yumpu.com/xx/document/view/52339780/42-/287″ title=»И. М. Эрлихсон. “Тре»>И. М. Эрлихсон. “Тре
И. М. Эрлихсон. “Тре
И. М. Эрлихсон. “Тре
И. М. Эрлихсон. “Тре
И. М. Эрлихсон. “Тре
И. М. Эрлихсон. “Тре
И. М. Эрлихсон. “Тре
 yumpu.com/xx/document/view/52339780/42-/301″ title=»И. М. Эрлихсон. “Тре»>И. М. Эрлихсон. “Тре
yumpu.com/xx/document/view/52339780/42-/301″ title=»И. М. Эрлихсон. “Тре»>И. М. Эрлихсон. “Тре
И. М. Эрлихсон. “Тре
И. И. Варьяш. Истори
И. И. Варьяш. Истори
И. И. Варьяш. Истори
И. И. Варьяш. Истори
И. И. Варьяш. Истори
 yumpu.com/xx/document/view/52339780/42-/315″ title=»И. И. Варьяш. Истори»>И. И. Варьяш. Истори
yumpu.com/xx/document/view/52339780/42-/315″ title=»И. И. Варьяш. Истори»>И. И. Варьяш. Истори
И. И. Варьяш. Истори
И. И. Варьяш. Истори
В. В. Гурин. Моральн
В. В. Гурин. Моральн
В. В. Гурин. Моральн
Д. В. Кирюхин. “Исто
 yumpu.com/xx/document/view/52339780/42-/329″ title=»Д. В. Кирюхин. “Исто»>Д. В. Кирюхин. “Исто
yumpu.com/xx/document/view/52339780/42-/329″ title=»Д. В. Кирюхин. “Исто»>Д. В. Кирюхин. “Исто
Д. В. Кирюхин. “Исто
Л. В. КЛЕПИКОВАОБРА
Л. В. Клепикова. Обр
Л. В. Клепикова. Обр
Л. В. Клепикова. Обр
Л. В. Клепикова. Обр
 yumpu.com/xx/document/view/52339780/42-/343″ title=»Я. Ю. Мукосеева. Ура»>Я. Ю. Мукосеева. Ура
yumpu.com/xx/document/view/52339780/42-/343″ title=»Я. Ю. Мукосеева. Ура»>Я. Ю. Мукосеева. Ура
Я. Ю. Мукосеева. Ура
Я. Ю. Мукосеева. Ура
Я. Ю. Мукосеева. Ура
А. В. ГНОЕВЫХТЕМА РЕ
А. В. Гноевых. Тема р
А. В. Гноевых. Тема р
 yumpu.com/xx/document/view/52339780/42-/357″ title=»А. В. Гноевых. Тема р»>А. В. Гноевых. Тема р
yumpu.com/xx/document/view/52339780/42-/357″ title=»А. В. Гноевых. Тема р»>А. В. Гноевых. Тема р
Д. С. ПЛАТОНОВФЕВРА
Д. С. Платонов. Февр
Д. С. Платонов. Февр
Д. С. Платонов. Февр
Д. С. Платонов. Февр
Читая книги… 373пли
 yumpu.com/xx/document/view/52339780/42-/371″ title=»Читая книги… 375дух»>Читая книги… 375дух
yumpu.com/xx/document/view/52339780/42-/371″ title=»Читая книги… 375дух»>Читая книги… 375дух
Читая книги… 377- ок
Читая книги… 379был
Читая книги… 381вну
Читая книги… 383ние
Читая книги… 385еще
Читая книги… 387Нап
 yumpu.com/xx/document/view/52339780/42-/385″ title=»Читая книги… 389Нак»>Читая книги… 389Нак
yumpu.com/xx/document/view/52339780/42-/385″ title=»Читая книги… 389Нак»>Читая книги… 389Нак
SUMMARIESALLAN MEGILL. For a global
Summaries 393Keywords: monastic lib
Summaries 395fried Wilhelm von Leib
397In the space of gender historyV.
399В пространстве ге
Каталог однонуклеотидных изменений, отличающих современных людей от архаичных гомининов.

Паабо С. Состояние человека — молекулярный подход. Cell 157 , 216–226 (2014).
PubMed Статья CAS PubMed Central Google Scholar
Kuhlwilm, M. et al. . Древние гены перетекают от ранних современных людей к восточным неандертальцам. Природа 530 , 429–433 (2016).
ADS CAS PubMed PubMed Central Статья Google Scholar
Шлебуш, К. М. и др. . Геномная изменчивость в семи группах кхоэ-сан раскрывает адаптацию и сложную африканскую историю. Наука (80-.). 338 , 374 LP – 379 (2012).
ADS Статья CAS Google Scholar
Hublin, J.-J. и др. .Новые окаменелости из Джебель-Ирхуда, Марокко и панафриканского происхождения Homo sapiens. Природа 546 , 289 (2017).
ADS CAS PubMed Статья PubMed Central Google Scholar
Шлебуш, К. М. и др. . Древние геномы южной части Африки оценивают расхождение современного человека от 350 000 до 260 000 лет назад. Наука (80-.) (2017).
Скоглунд, П. и др. . Реконструкция доисторической структуры населения Африки. Cell 171 , 59–71.e21 (2017).
CAS PubMed PubMed Central Статья Google Scholar
Rabett, R.J. Успех неудачного расселения Homo sapiens из Африки в Азию. Nat. Ecol. Evol. 2 , 212–219 (2018).
PubMed Статья PubMed Central Google Scholar
Posth, C. и др. . Глубоко дивергентный архаический митохондриальный геном обеспечивает более низкую временную границу для потока африканских генов в неандертальцев. 8 , 16046 (2017).
8 , 16046 (2017).
Hublin, J.-J., Neubauer, S. & Gunz, P. Онтогенез мозга и история жизни плейстоценовых гомининов. Philos. Пер. R. Soc. B Biol. Sci . 370 (2015).
Гунц, П., Нойбауэр, С., Мориль, Б. и Хаблин, Дж. Дж. Развитие мозга после рождения отличается у неандертальцев и современных людей. Curr. Биол. 20 , 921–922 (2010).
Артикул CAS Google Scholar
Neubauer, S., Hublin, J.-J. И Гунц, П. Эволюция современной формы человеческого мозга. Sci. Adv . 4 (2018).
Gunz, P. et al. . Интрогрессия неандертальцев проливает свет на эндокраниальную глобулярность современного человека. Curr. Биол. 29 , 120–127.e5 (2019).
CAS PubMed Статья PubMed Central Google Scholar
Тринкаус, Э. и Хауэллс, У. В. Неандертальцы. Sci. Являюсь. 241 , 118–133 (1979).
В. Неандертальцы. Sci. Являюсь. 241 , 118–133 (1979).
Артикул Google Scholar
Шенеманн, П. Т. Масштабирование размера мозга и состав тела у млекопитающих. Мозг. Behav. Evol. 63 , 47–60 (2004).
PubMed Статья PubMed Central Google Scholar
Gunz, P. et al. . Уникальный современный образец эндокраниального развития человека. Выводы из новой черепной реконструкции новорожденного неандертальца из Мезмайской. J. Hum. Evol. 62 , 300–313 (2012).
PubMed Статья PubMed Central Google Scholar
Boeckx, C. Голова, готовая к работе с языком: эволюционные соображения. Психон. Бык. Ред. 24 , 194–199 (2017).
PubMed Статья PubMed Central Google Scholar

Хоффманн, Д. Л., Ангелуччи, Д. Э., Вильяверде, В., Сапата, Дж. И Зильхао, Дж. Символическое использование морских раковин и минеральных пигментов иберийскими неандертальцами 115 000 лет назад. Sci. Adv . 4 (2018).
Винн Т., Оверманн К. и Кулидж Ф. Ложная дихотомия: опровержение утверждения о неразличимости неандертальцев. J. Anthropol. Sci. = Riv. ди Антрополь. JASS 94 , 201–221 (2016).
Google Scholar
Мейер, М. и др. . Последовательность генома с высоким охватом от архаичного денисовского индивида. Наука 338 , 222–226 (2012).
ADS CAS PubMed PubMed Central Статья Google Scholar
Prüfer, K. et al . Полная последовательность генома неандертальца из Горного Алтая. Природа 505 , 43–9 (2014).
ADS PubMed PubMed Central Статья CAS Google Scholar

Prüfer, K. et al. . Геном неандертальца с высоким охватом из пещеры Виндия в Хорватии. Наука (80-.) (2017).
Auton, A. et al. .Глобальный справочник по генетическим вариациям человека. Природа 526 , 68–74 (2015).
ADS PubMed PubMed Central Статья CAS Google Scholar
Consortium, T. C. S. и A. Исходная последовательность генома шимпанзе и сравнение с геномом человека. Природа 437 , 69 (2005).
Артикул CAS Google Scholar
Ju, X.-C. и др. . Гоминоид-специфический ген TBC1D3 способствует генерации базальных нейральных предшественников и индуцирует кортикальный фолдинг у мышей. Элиф 5 , e18197 (2016).
PubMed PubMed Central Статья Google Scholar

Флорио, М. и др. . Специфический для человека ген ARHGAP11B способствует базовой амплификации предшественников и расширению неокортекса. Наука 347 , 1465–70 (2015).
ADS CAS PubMed Статья Google Scholar
Дюма, Л. Дж. и др. . Число копий домена DUF1220, влияющее на патологию и эволюцию размера мозга человека. г. J. Hum. Genet. 91 , 444–454 (2012).
CAS PubMed PubMed Central Статья Google Scholar
Деннис, М. Ю. и др. .Эволюция человеческих нейронных генов SRGAP2 путем неполной сегментарной дупликации. Cell 149 , 912–922 (2012).
CAS PubMed PubMed Central Статья Google Scholar
Сузуки, И. К. и др. . Специфические для человека гены NOTCh3NL расширяют корковый нейрогенез за счет регуляции Delta / Notch. Ячейка 173 , 1370–1384.e16 (2018).
Ячейка 173 , 1370–1384.e16 (2018).
CAS PubMed PubMed Central Статья Google Scholar
Маклин, К. Ю. и др. . Специфическая для человека потеря регуляторной ДНК и эволюция специфических для человека черт. Природа 471 , 216–219 (2011).
ADS CAS PubMed PubMed Central Статья Google Scholar
Чинталапати, М., Даннеманн, М. и Прюфер, К. Использование генома неандертальца для изучения эволюции небольших вставок и делеций у современных людей. BMC Evol.Биол. 17 , 179 (2017).
PubMed PubMed Central Статья Google Scholar
Castellano, S. et al . Паттерны вариации кодирования в полных экзомах трех неандертальцев. Proc. Natl. Акад. Sci. США 111 , 6666–71 (2014).
ADS CAS PubMed Статья PubMed Central Google Scholar

Грин, Р. Э. и др. . Предварительная последовательность генома неандертальца. Наука (80-.). 328 , 710–722 (2010).
ADS CAS PubMed Central Статья Google Scholar
Пейрегне, С., Бойл, М. Дж., Даннеманн, М. и Прюфер, К. Обнаружение древнего положительного отбора у людей с использованием расширенной сортировки по родословным. Genome Res. 27 , 1563–1572 (2017).
PubMed PubMed Central Статья CAS Google Scholar
Расимо, Ф. Тестирование древней селекции с использованием межпопуляционной дифференциации частоты аллелей. Genetics 202 , 733 LP – 750 (2016).
Артикул CAS Google Scholar
Somel, M., Liu, X. & Khaitovich, P. Эволюция человеческого мозга: транскрипты, метаболиты и их регуляторы. Nat. Rev. Neurosci. 14 , 1–16 (2013).
Rev. Neurosci. 14 , 1–16 (2013).
Артикул CAS Google Scholar
Чжоу, Х. и др. . Хронологический атлас естественного отбора в геноме человека за последние полмиллиона лет. bioRxiv (2015).
Sankararaman, S. et al. . Геномный ландшафт неандертальцев у современных людей. Природа 507 , 354–357 (2014).
ADS CAS PubMed PubMed Central Статья Google Scholar
Верно, Б. и Эйки, Дж. М. Возрождение выживших неандертальских линий из геномов современного человека. Наука (80-.). 343 , 1017–1021 (2014).
ADS CAS Статья Google Scholar
Vernot, B. et al. . Выделение ДНК неандертальцев и денисовцев из геномов меланезийских особей. Наука (80-.). 352 , 235–239 (2016).
ADS CAS Статья Google Scholar
Санкарараман С., Маллик С., Паттерсон Н. и Райх Д. Объединенный ландшафт денисовских и неандертальских предков у современных людей. Curr. Биол. 26 , 1241–1247 (2016).
CAS PubMed PubMed Central Статья Google Scholar
Маккой Р.С., Уэйкфилд Дж. И Эйки Дж. М. Влияние интрогрессированных неандертальцами последовательностей на ландшафт экспрессии генов человека. Ячейка 168 , 916–927.e12 (2017).
CAS PubMed PubMed Central Статья Google Scholar
Винн Т. и Кулидж Ф. Л. Эксперт в области разума неандертальцев. J. Hum. Evol. 46 , 467–487 (2004).
PubMed Статья Google Scholar
Langergraber, K. E. et al. . Время генерации у диких шимпанзе и горилл предполагает более раннее время расхождения в эволюции человекообразных обезьян и людей. Proc. … 109 , 15716–15721 (2012).
et al. . Время генерации у диких шимпанзе и горилл предполагает более раннее время расхождения в эволюции человекообразных обезьян и людей. Proc. … 109 , 15716–15721 (2012).
CAS Google Scholar
Варки А. и Альтейде Т. К. Сравнение геномов человека и шимпанзе: поиск иголок в стоге сена. Genome Res. 15 , 1746–1758 (2005).
CAS PubMed Статья Google Scholar
О’Блесс, М., Сирлз, В. Б., Варки, А., Гагнё, П. и Сикела, Дж. М. Эволюция генетических и геномных особенностей, уникальных для человеческого происхождения. Nat. Преподобный Жене. 13 , 853–866 (2012).
PubMed PubMed Central Статья CAS Google Scholar
Bufill, E., Blesa, R. & Augusti, J. Болезнь Альцгеймера: эволюционный подход. J. Anthropol. Sci. = Riv. ди Антрополь. JASS 91 , 135–157 (2013).
= Riv. ди Антрополь. JASS 91 , 135–157 (2013).
Google Scholar
Буфилл, Э., Агусти, Дж. И Блеса, Р. Возвращение к неотении человека: случай синаптической пластичности. г. J. Hum. Биол. 23 , 729–739 (2011).
PubMed Статья PubMed Central Google Scholar
Брунер, Э. и Джейкобс, Х. И. Л. Болезнь Альцгеймера: обратная сторона высокоразвитой теменной доли? Дж.Альцгеймера. Дис. 35 , 227–240 (2013).
CAS PubMed Статья PubMed Central Google Scholar
Граттен, Дж. И Висшер, П. М. Генетическая плейотропия сложных признаков и заболеваний: значение для геномной медицины. Genome Med. 8 , 78 (2016).
PubMed PubMed Central Статья CAS Google Scholar

Джандоменико, С. Л. и Ланкастер, М. А. Исследование эволюции и развития человеческого мозга с помощью органоидов. Curr. Opin. Cell Biol. 44 , 36–43 (2017).
CAS PubMed Статья PubMed Central Google Scholar
Кэмп, Дж. Г. и др. . Органоиды головного мозга человека повторяют программы экспрессии генов развития неокортекса плода. Proc. Natl. Акад. Sci. США 112 , 15672–7 (2015).
ADS CAS PubMed Статья PubMed Central Google Scholar
Кэмп, Дж. Г. и Трейтлейн, Б. Человеческая органика: новый подход к пониманию человеческого развития с использованием транскриптомики одиночных клеток. Девелопмент 144 , 1584 LP – 1587 (2017).
Артикул CAS Google Scholar
Пыльца А.А. и др. .Установление церебральных органоидов как моделей эволюции мозга человека. Ячейка 176 , 743–756.e17 (2019).
Ячейка 176 , 743–756.e17 (2019).
CAS PubMed Статья PubMed Central Google Scholar
Enard, W. et al. . Гуманизированная версия Foxp2 влияет на контуры кортико-базальных ганглиев у мышей. Cell 137 , 961–971 (2009).
CAS PubMed Статья PubMed Central Google Scholar
MacArthur, J. et al. . Новый Каталог опубликованных полногеномных ассоциативных исследований NHGRI-EBI (Каталог GWAS). Nucleic Acids Res. 45 , D896 – D901 (2017).
CAS PubMed Статья PubMed Central Google Scholar
Рэй, Г. А. Эволюционное значение цис-регуляторных мутаций. Nat. Преподобный Жене. 8 , 206 (2007).
CAS PubMed Статья PubMed Central Google Scholar

Чавла, К., Трипати, С., Томмесен, Л., Легрейд, А. и Койпер, М. TFcheckpoint: тщательно подобранный сборник факторов транскрипции специфической ДНК-связывающей РНК-полимеразы II. Биоинформатика 29 , 2519–2520 (2013).
CAS PubMed Статья PubMed Central Google Scholar
Гроте, С., Прюфер, К., Келсо, Дж. И Даннеманн, М. ABAEnrichment: пакет R для тестирования обогащения экспрессии набора генов в мозге взрослого и развивающегося человека. Биоинформатика 32 , 3201–3203 (2016).
CAS PubMed PubMed Central Статья Google Scholar
Миллер Дж. А. и др. . Транскрипционный ландшафт пренатального мозга человека. Природа 508 , 199–206 (2014).
ADS CAS PubMed PubMed Central Статья Google Scholar

де Бур Б., Томпсон Б., Равиньяни А. и Беккс К. Эволюционная динамика не мотивирует единомутантную теорию человеческого языка. bioRxiv 517029 , https://doi.org/10.1101/517029 (2019).
Омер Джавед, А. и др. . Моделирование микроцефалии мутации кинетохор выявляет специфический для мозга фенотип. Cell Rep. 25 , 368–382.e5 (2018).
CAS PubMed Статья PubMed Central Google Scholar
О’Реган, Л. и Фрай, А. М. Протеинкиназы Nek6 и Nek7 необходимы для надежного формирования митотического веретена и цитокинеза. Мол. Клетка. Биол. 29 , 3975–3990 (2009).
PubMed PubMed Central Статья CAS Google Scholar
Торрес, Дж. З. и др. . Кинезин STARD9 / Kif16a ассоциирует с митотическими микротрубочками и регулирует сборку полюсов веретена. Cell 147 , 1309–1323 (2017).
Артикул CAS Google Scholar
Барр, А. Р., Килмартин, Дж. В. и Гергели, Ф. CDK5RAP2 функционирует в центросоме для прикрепления полюса веретена и ответа на повреждение ДНК. J. Cell Biol. 189 , 23 ЛП – 39 (2010).
Артикул CAS Google Scholar
Пуга А., Ся Ю. и Эльферинк С. Роль арилуглеводородного рецептора в регуляции клеточного цикла. Chem. Биол. Взаимодействовать. 141 , 117–130 (2002).
CAS PubMed Статья PubMed Central Google Scholar
Бадер, Дж. Р. и др. . Поло-подобная киназа1 необходима для рекрутирования динеина в кинетохоры во время митоза. J. Biol. Chem. 286 , 20769–20777 (2011).
CAS PubMed PubMed Central Статья Google Scholar

Zachos, G. et al. . Chk1 требуется для функции контрольной точки шпинделя. Dev. Ячейка 12 , 247–260 (2017).
Артикул CAS Google Scholar
Йошида М. и Азума Ю. Механизмы SUMOилирования топоизомеразы II при сегрегации хромосом. Cell Cycle 15 , 3151–3152 (2016).
CAS PubMed PubMed Central Статья Google Scholar
Фиц, С. А. и Хаттнер, В. Б. Расширение корковых предшественников, самообновление и нейрогенез — поляризованная перспектива. Curr. Opin. Neurobiol. 21 , 23–35 (2011).
CAS PubMed Статья PubMed Central Google Scholar
Megraw, T. L., Sharkey, J. T. и Nowakowski, R. S. Cdk5rap2 выявляет центросомный корень синдромов микроцефалии. Trends Cell Biol. 21 , 470–480 (2011).
CAS PubMed PubMed Central Статья Google Scholar
Гупта, Г. Д. и др. . Динамический ландшафт взаимодействия белков на границе раздела центросома-ресничка человека. Cell 163 , 1484–1499 (2015).
CAS PubMed PubMed Central Статья Google Scholar
Li, F.-F. и др. .Идентификация двух новых критических мутаций в гене PCNT, приводящих к микроцефальному остеодиспластическому первичному карликовому типу II, связанному с множественными внутричерепными аневризмами. Metab. Brain Dis. 30 , 1387–1394 (2015).
CAS PubMed Статья PubMed Central Google Scholar
Ричи, К., Уотсон, Л. А., Дэвидсон, Б., Цзян, Ю. и Берубе, Н. Г. ATRX требуется для поддержания пула клеток-предшественников в эмбриональном мозге мыши. Biol. Открыть 3 , 1158 LP – 1163 (2014).
Biol. Открыть 3 , 1158 LP – 1163 (2014).
Артикул Google Scholar
Генин А. и др. . Ген CASC5 кинетохорной сети KMN мутировал при первичной микроцефалии. Hum. Мол. Genet. 21 , 5306–5317 (2012).
CAS PubMed Статья PubMed Central Google Scholar
Montgomery, S.Х., Капеллини И., Вендитти К., Бартон Р. А. и Манди Н. И. Адаптивная эволюция четырех генов микроцефалии и эволюция размера мозга у антропоидных приматов. Мол. Биол. Evol. 28 , 625–638 (2011).
CAS PubMed Статья PubMed Central Google Scholar
Shi, L. et al. . Региональный отбор гена CASC5, регулирующего размер мозга, позволяет по-новому взглянуть на эволюцию человеческого мозга. Hum. Genet. 136 , 193–204 (2017).
CAS PubMed Статья PubMed Central Google Scholar

Chen, Y., Huang, W.-C., Séjourné, J., Clipperton-Allen, AE & Page, DT Pten Мутации изменяют траекторию роста мозга и распределение типов клеток за счет повышения β- Катенин сигнализация. J. Neurosci. 35 , 10252 LP – 10267 (2015).
Артикул CAS Google Scholar
Li, Y. и др. . Индукция расширения и складывания органоидов головного мозга человека. Cell Stem Cell 20 , 385–396.e3 (2017).
PubMed Статья CAS PubMed Central Google Scholar
Kuwahara, A. et al. . Tcf3 подавляет передачу сигналов Wnt – β-катенин и поддерживает популяцию нервных стволовых клеток во время развития неокортекса. PLoS One 9 , e94408 (2014).
ADS PubMed PubMed Central Статья CAS Google Scholar
Мирзаа Г. М. и др. . Мутации de novo CCND2, приводящие к стабилизации циклина D2, вызывают синдром мегалэнцефалии-полимикрогирии-полидактилии-гидроцефалии. Nat. Genet. 46 , 510 (2014).
М. и др. . Мутации de novo CCND2, приводящие к стабилизации циклина D2, вызывают синдром мегалэнцефалии-полимикрогирии-полидактилии-гидроцефалии. Nat. Genet. 46 , 510 (2014).
CAS PubMed PubMed Central Статья Google Scholar
Джамшир, А. и др. . Расширенный спектр мутаций гена GLI3 подтверждает корреляции генотип-фенотип. J. Appl. Genet. 53 , 415–422 (2012).
CAS PubMed PubMed Central Статья Google Scholar
Wang, C., Liang, C.-C., Bian, Z. C., Zhu, Y. & Guan, J.-L. FIP200 необходим для поддержания и дифференцировки постнатальных нервных стволовых клеток. Nat.Neurosci. 16 , 532 (2013).
PubMed PubMed Central Статья CAS Google Scholar
Weyer, S. & Pääbo, S. Функциональные анализы сайтов связывания транскрипционных факторов, которые различаются между современными и архаичными людьми. Мол. Биол. Evol. 33 , 316–322 (2016).
Мол. Биол. Evol. 33 , 316–322 (2016).
CAS PubMed Статья PubMed Central Google Scholar
Кочияма, Т. и др. . Реконструкция мозга неандертальца с использованием вычислительной анатомии. Sci. Отчет 8 , 6296 (2018).
ADS PubMed PubMed Central Статья CAS Google Scholar
Миллер, И. Ф., Бартон, Р. А. и Нанн, К. Л. Количественная уникальность эволюции человеческого мозга, выявленная с помощью филогенетического сравнительного анализа. Элиф 8 , e41250 (2019).
PubMed PubMed Central Статья Google Scholar
Бланк, М.С. и др. . Множественные программы развития изменяются из-за потери Zic1 и Zic4 , что вызывает патогенез мозжечка у порока Денди-Уокера. Девелопмент 138 , 1207 LP – 1216 (2011).
Артикул CAS Google Scholar
Cheng, Y.-Z. и др. . Изучение паттернов эмбриональной экспрессии и эволюции генов AHI1 и CEP290, связанных с синдромом Жубера. PLoS One 7 , e44975 (2012).
ADS CAS PubMed PubMed Central Статья Google Scholar
Hoshino, J., Aruga, J., Ishiguro, A. & Mikoshiba, K. Dorz1, новый ген, экспрессирующийся в дифференцирующихся нейронах гранул мозжечка, подавляется у Zic1-дефицитных мышей. Мол. Brain Res. 120 , 57–64 (2003).
CAS PubMed Статья PubMed Central Google Scholar
Pfenning, A. R. et al. . Конвергентные транскрипционные специализации в мозге людей и певчих птиц. Наука (80-.) . 346 (2014).
Wang, R. et al. . Конвергентная дифференциальная регуляция генов, управляющих аксонами SLIT-ROBO в мозге обучающихся вокалу. J. Comp. Neurol. 523 , 892–906 (2015).
J. Comp. Neurol. 523 , 892–906 (2015).
CAS PubMed Статья Google Scholar
Kuhlwilm, M. Эволюция FOXP2 в свете примесей. Curr. Opin. Behav. Sci . 21 (2018).
Артикул Google Scholar
Фишер С.Э. Генетика человека: эволюционирующая история FOXP2. Curr.Биол. 29 , R65 – R67 (2019).
CAS PubMed Статья PubMed Central Google Scholar
Людерс, Дж. Цитоскелет микротрубочек , https://doi.org/10.1007/978-3-7091-1903-7 (Springer Vienna, 2016).
Google Scholar
McVicker, D. P. et al. . Транспорт пары кинезин-груз по микротрубочкам в дендритные шипы, претерпевающие синаптическую пластичность. Nat. Commun. 7 , 12741 (2016).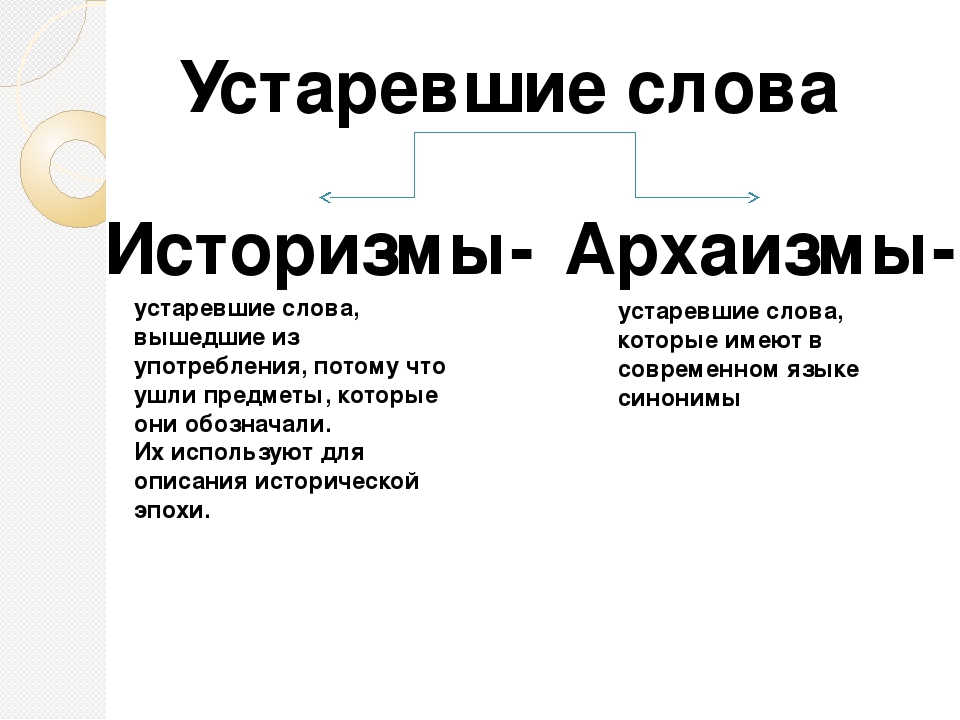
ADS CAS PubMed PubMed Central Статья Google Scholar
Ахмад, Ф. Дж., Ю, В., МакНалли, Ф. Дж. И Баас, П. В. Существенная роль катанина в разделении микротрубочек в нейроне. J. Cell Biol. 145 , 305 LP – 315 (1999).
Артикул Google Scholar
Карабай, А., Ю., В., Соловска, Дж. М., Бэрд, Д. Х. и Баас, П. В. Рост аксонов чувствителен к уровням катанина, белка, разделяющего микротрубочки. J. Neurosci. 24 , 5778 LP – 5788 (2004).
Артикул CAS Google Scholar
Кевенаар, Дж. Т. и др. . Кинезин-связывающий белок контролирует динамику микротрубочек и перемещение грузов путем регулирования моторной активности кинезина. Curr.Биол. 26 , 849–861 (2017).
Артикул CAS Google Scholar

Wu, Y. et al. . Изоформа Versican V1 вызывает дифференцировку нейронов и способствует росту нейритов. Мол. Биол. Ячейка 15 , 2093–2104 (2004).
CAS PubMed PubMed Central Статья Google Scholar
Куберос, Х. и др. . Роль киназ LIM в функции и дисфункции центральной нервной системы. FEBS Lett. 589 , 3795–3806 (2015).
CAS PubMed Статья PubMed Central Google Scholar
Koser, D. E. et al. . Механочувствительность имеет решающее значение для роста аксонов в развивающемся головном мозге. Nat. Neurosci. 19 , 1592 (2016).
CAS PubMed PubMed Central Статья Google Scholar
Дженсен, К. Б. и др. . Nova-1 регулирует альтернативное сплайсинг, специфичное для нейронов, и имеет важное значение для жизнеспособности нейронов. Нейрон 25 , 359–371 (2000).
Нейрон 25 , 359–371 (2000).
CAS PubMed Статья Google Scholar
Ratti, A. et al. . Посттранскрипционная регуляция нейроонкологического вентрального антигена 1 нейрональными РНК-связывающими белками ELAV. J. Biol. Chem ., Https: // doi.org / 10.1074 / jbc.M706082200 (2008 г.).
CAS PubMed Статья Google Scholar
Takemoto, M. et al . Ламинарная и ареальная экспрессия Unc5d и ее роль в выживании корковых клеток. Cereb. Cortex 21 , 1925–1934 (2011).
PubMed Статья Google Scholar
Яп, К. К. и др. . Даблкортин (DCX) опосредует эндоцитоз нейрофасцина независимо от связывания микротрубочек. J. Neurosci. 32 , 7439 LP – 7453 (2012).
Артикул CAS Google Scholar

Verheijen, M.H.G. et al. . SCAP необходим для своевременного и правильного синтеза миелиновой мембраны. Proc. Natl. Акад. Sci. 106 , 21383–21388 (2009).
ADS CAS PubMed Статья Google Scholar
Мензис Ф. М., Флеминг А. и Рубинштейн Д. С. Нарушение аутофагии и нейродегенеративные заболевания. Nat. Rev. Neurosci. 16 , 345 (2015).
CAS PubMed Статья Google Scholar
Jurecka, A., Jurkiewicz, E. & Tylki-Szymanska, A. Магнитно-резонансная томография мозга при дефиците аденилосукцинатлиазы: отчет о семи случаях и обзор литературы. евро.J. Pediatr. 171 , 131–138 (2012).
PubMed Статья Google Scholar
Widagdo, J., Fang, H., Jang, S. E. и Anggono, V. PACSIN1 регулирует динамику передачи рецептора AMPA. Sci. Отчет 6 , 31070 (2016).
Sci. Отчет 6 , 31070 (2016).
ADS CAS PubMed PubMed Central Статья Google Scholar
Скейде, М. А. и Фридеричи, А. Д. Онтогенез корковой языковой сети. Nat. Rev. Neurosci. 17 , 323 (2016).
CAS PubMed Статья Google Scholar
Гроссе Висманн, К., Шрайбер, Дж., Сингер, Т., Стейнбейс, Н. и Фридеричи, А. Д. Созревание белого вещества связано с возникновением теории разума в раннем детстве. Nat. Commun. 8 , 14692 (2017).
ADS CAS PubMed PubMed Central Статья Google Scholar
Марс, Р. Б., Эйхерт, Н., Джбабди, С., Верхаген, Л. и Рашворт, М. Ф. С. Связь и поиск специализаций в способном к языку мозге. Curr. Opin. Behav. Sci. 21 , 19–26 (2018).
Артикул Google Scholar

Ардеш, Д.J. и др. . Эволюционное расширение связи между областями мультимодальных ассоциаций в мозге человека по сравнению с шимпанзе. Proc. Natl. Акад. Sci. 116 , 7101 LP – 7106 (2019).
Артикул CAS Google Scholar
Dubois, J. et al. . Изучение ранней организации и созревания языковых путей в мозге младенца. Cereb. Cortex 26 , 2283–2298 (2016).
PubMed Статья PubMed Central Google Scholar
Абельсон, Дж. Ф. и др. . Варианты последовательности в SLITRK1 связаны с синдромом Туретта. Наука . 310 , 317 ЛП – 320 (2005 г.).
ADS Статья CAS Google Scholar
Jelen, N., Ule, J. & ivin, M. Холинергическая регуляция мРНК новой полосатого тела. Неврология 169 , 619–627 (2010).
CAS PubMed Статья PubMed Central Google Scholar
Алсио, Дж. М., Тарчини, Б., Кайуэтт, М. и Ливси, Ф. Дж. Икарос способствует формированию судьбы ранних нейронов в коре головного мозга. Proc. Natl. Акад. Sci. 110 , E716 – E725 (2013).
ADS PubMed Статья PubMed Central Google Scholar
Поповиченко Т. и др. . РНК-связывающий белок HuR определяет дифференциальную трансляцию ассоциированных с аутизмом членов подсемейства FoxP в развивающемся неокортексе. Sci. Отчет 6 , 28998 (2016).
ADS CAS PubMed PubMed Central Статья Google Scholar
Конопка, Г. и др. . Специфичные для человека транскрипционные сети в головном мозге. Нейрон 75 , 601–617 (2012).
CAS PubMed PubMed Central Статья Google Scholar

Sreedharan, S. et al. . Рецептор, связанный с G-белком, Gpr153 имеет общее эволюционное происхождение с Gpr162 и высоко экспрессируется в центральных областях, включая таламус, мозжечок и дугообразное ядро. FEBS J. 278 , 4881–4894 (2011).
CAS PubMed Статья PubMed Central Google Scholar
Takata, A. et al. . Интегративный анализ мутаций De Novo обеспечивает более глубокое биологическое понимание расстройства аутистического спектра. Cell Rep. 22 , 734–747 (2018).
CAS PubMed Статья PubMed Central Google Scholar
Фон, Э. А. и др. . Аденилосукцинатлиаза (ADSL) и детский аутизм: отсутствие точечной мутации, о которой ранее сообщалось. г. Дж.Med. Genet. 60 , 554–557 (1995).
CAS PubMed Статья PubMed Central Google Scholar

Racimo, F., Kuhlwilm, M. & Slatkin, M. Тест для древних выборочных обследований и его применение к кандидатным участкам у современных людей. Мол. Биол. Evol. 31 , 3344–3358 (2014).
CAS PubMed PubMed Central Статья Google Scholar
Дедиу Д. и Левинсон С. С. О древности языка: переосмысление языковых способностей неандертальцев и их последствия. Фронт. Psychol. 4 , 1–17 (2013).
Артикул Google Scholar
Dediu, D. & Levinson, S. C. Еще раз о неандертальском языке: не только мы. Curr. Opin. Behav. Sci. 21 , 49–55 (2018).
Артикул Google Scholar
Fu, Q. и др. . Ранний современный человек из Румынии, недавний предок — неандерталец. Природа 524 , 216–219 (2015).
ADS CAS PubMed PubMed Central Статья Google Scholar

Lei, H. et al. . Пути наведения аксонов служили обычными мишенями для эволюции речи / языка человека и связанных с ним расстройств. Brain Lang. 174 , 1–8 (2017).
PubMed Статья PubMed Central Google Scholar
Jeong, J.-W., Sundaram, S., Behen, M. E. & Chugani, H. T. Взаимосвязь между генотипом и морфологией дугообразного пучка у шести маленьких детей с глобальной задержкой развития: предварительное исследование DTI. J. Magn. Резон. Imaging 44 , 1504–1512 (2016).
PubMed PubMed Central Статья Google Scholar
Беккс, К. и Бенитес-Буррако, А. Глобальность и языковая готовность: создание новых прогнозов путем расширения набора представляющих интерес генов. Границы в психологии 5 , 1324 (2014).
PubMed PubMed Central Google Scholar

Bastir, M. et al. . Эволюция основания мозга у людей с высокой энцефализацией. Nat. Commun. 2 , 588 (2011).
PubMed Статья CAS PubMed Central Google Scholar
Брайант, К. Л. и Прейс, Т. М. Сравнительный взгляд на височную долю человека BT — Цифровые эндокасты: от черепа до мозга. в (ред. Брунер, Э., Огихара, Н. и Танабе, Х. К.) 239–258, https://doi.org/10.1007/978-4-431-56582-6_16 (Springer Japan, 2018).
Google Scholar
Гохман Д. и др. . Обширные регуляторные изменения в генах, влияющих на анатомию голоса и лица, отделяют современных людей от архаичных. bioRxiv (2017).
Белик М. и Браун С. Происхождение голосового мозга у человека. Neurosci. Biobehav. Ред. 77 , 177–193 (2017).
PubMed Статья PubMed Central Google Scholar

Адхикари, К. и др. . Полногеномное сканирование ассоциаций вовлекает DCHS2, RUNX2, GLI3, PAX1 и EDAR в изменение лица человека. Nat. Commun. 7 , 11616 (2016).
ADS CAS PubMed PubMed Central Статья Google Scholar
Ким, Дж. Х. и Ким, Н. Регулирование NFATc1 в дифференцировке остеокластов. J Bone Metab 21 , 233–241 (2014).
ADS PubMed PubMed Central Статья Google Scholar
Cai, H. & Liu, A. Spop способствует развитию скелета и гомеостазу, положительно регулируя передачу сигналов Ihh. Proc. Natl. Акад. Sci. 113 , 14751–14756 (2016).
CAS PubMed Статья PubMed Central Google Scholar
Zhang, Y. et al. . Существенная роль рецептора домена дискоидина 2 (DDR2) в дифференцировке остеобластов и созревании хондроцитов посредством модуляции активации Runx2. J. Bone Miner. Res. 26 , 604–617 (2011).
J. Bone Miner. Res. 26 , 604–617 (2011).
CAS PubMed Статья PubMed Central Google Scholar
Zhang, X. et al. . Дефекты, подобные кледокраниодисплазии свода черепа, с дефицитом Nell-1, вызванным ENU. J. Craniofac. Сургут . 23 (2012).
CAS PubMed Статья PubMed Central Google Scholar
Мончини, С. и др. . Мутация ATRX у двух взрослых братьев с неспецифической умеренной умственной отсталостью, выявленная с помощью секвенирования экзома. Мета-ген 1 , 102–108 (2013).
CAS PubMed PubMed Central Статья Google Scholar
Oh, J.-E. и др. . PlexinA2 опосредует дифференцировку остеобластов посредством регуляции Runx2. J. Bone Miner. Res. 27 , 552–562 (2012).
CAS PubMed Статья PubMed Central Google Scholar

Квон, Э. К. и др. . Роль Эллис-Ван Кревельда 2 ( EVC2 ) у мышей во время развития костей черепа. Анат. Рек. 301 , 46–55 (2018).
CAS Статья Google Scholar
Gullard, A. et al. . Локализация MEPE в черепно-лицевом комплексе и функция в формировании зубного дентина. J. Histochem. Cytochem. 64 , 224–236 (2016).
CAS PubMed PubMed Central Статья Google Scholar
Tashima, T., Nagatoishi, S., Sagara, H., Ohnuma, S. & Tsumoto, K. Остеомодулин регулирует диаметр и изменяет форму коллагеновых фибрилл. Biochem. Биофиз. Res. Commun. 463 , 292–296 (2015).
CAS PubMed Статья PubMed Central Google Scholar
Тевес, М.Э. и др. . Дефицит Spag17 приводит к порокам развития скелета и костным аномалиям. PLoS One 10 , e0125936 (2015).
PLoS One 10 , e0125936 (2015).
PubMed PubMed Central Статья CAS Google Scholar
Lacruz, R. S. et al. . Онтогенез верхней челюсти у неандертальцев и их предков. Nat. Commun. 6 , 8996 (2015).
PubMed PubMed Central Статья CAS Google Scholar
Lacruz, R. S. et al. . Морфогенез лица древних европейцев. PLoS One 8 , e65199 (2013).
ADS CAS PubMed PubMed Central Статья Google Scholar
Стрингер К. Происхождение и эволюция Homo sapiens. Философия . Пер. R. Soc. B Biol. Sci . 371 (2016).
Сигел, Д. А., Хуанг, М.К. и Беккер, С. Ф. Инициирование эктопических дендритов: патогенез ЦНС как модель развития ЦНС. Внутр. J. Dev. Neurosci. 20 , 373–389 (2002).
20 , 373–389 (2002).
CAS PubMed Статья PubMed Central Google Scholar
Park, J.-A. И Ким, К. С. Паттерны экспрессии PRDM10 во время эмбрионального развития мыши. BMB Rep. 43 , 29–33 (2010).
CAS PubMed Статья PubMed Central Google Scholar
Джонс, Д. К. и др. . Несвязанность созревания пластинки роста и образования костей у мышей, лишенных как Schnurri-2, так и Schnurri-3. Proc. Natl. Акад. Sci. 107 , 8254–8258 (2010).
ADS CAS PubMed Статья PubMed Central Google Scholar
Имамура, К. и др. . Белок 3, связывающий энхансер вируса иммунодефицита человека типа 1, необходим для экспрессии связанного с аспарагином гликозилирования 2 в регуляции дифференцировки остеобластов и хондроцитов. J. Biol. Chem. 289 , 9865–9879 (2014).
J. Biol. Chem. 289 , 9865–9879 (2014).
CAS PubMed PubMed Central Статья Google Scholar
Ли, М. К. и др. . Полногеномное исследование ассоциации морфологии лица обнаруживает новые ассоциации с FREM1 и PARK2. PLoS One 12 , e0176566 (2017).
PubMed PubMed Central Статья CAS Google Scholar
Талбот, Дж. К. и др. . Морфогенез глотки требует fras1-itga8-зависимого эпителиально-мезенхимального взаимодействия. Dev. Биол. 416 , 136–148 (2016).
CAS PubMed PubMed Central Статья Google Scholar
Verzi, M. P. et al. . Фактор транскрипции MEF2C необходим для черепно-лицевого развития. Dev. Ячейка 12 , 645–652 (2007).
CAS PubMed PubMed Central Статья Google Scholar

Рак Ю. Неандерталец: новый взгляд на старое лицо. J. Hum. Evol. 15 , 151–164 (1986).
Артикул Google Scholar
Stelzer, S., Neubauer, S., Hublin, J.-J., Spoor, F. & Gunz, P. Морфологические тенденции в форме и размере аркад в среднем плейстоцене Homo. г. J. Phys. Антрополь. 168 , 70–91 (2019).
PubMed Статья PubMed Central Google Scholar
Somel, M. et al . Транскрипционная неотения в мозге человека. Proc. Natl. Акад. Sci. США 106 , 5743–8 (2009).
ADS CAS PubMed Статья PubMed Central Google Scholar
Лю, Х. и др. . Расширение коркового синаптического развития отличает человека от шимпанзе и макак. Genome Res. 22 , 611–622 (2012).
CAS PubMed PubMed Central Статья Google Scholar

Шервуд К. и Гомес-Роблес А. Пластичность мозга и эволюция человека. Annu. Преподобный Антрополь. 46 , 399–419 (2017).
Артикул Google Scholar
Беднарик, Г. Р. От человеческого прошлого к человеческому будущему. Гуманитарные науки 2 (2013).
Артикул Google Scholar
Hofman, M. a. Энцефализация у гоминидов: свидетельство модели пунктуации. Мозг. Behav. Evol ., Https://doi.org/10.1159/000121511 (1983).
CAS PubMed Статья PubMed Central Google Scholar
Понсе де Леон, М. С., Бьенвену, Т., Акадзава, Т., Цолликофер, К. П. Э. Развитие мозга у неандертальцев и современных людей сходно. Curr. Биол. 26 , R665 – R666 (2017).
Артикул CAS Google Scholar

Росас, А. и др. . Схема роста неандертальцев, реконструированная по скелету подростка из Эль-Сидрона (Испания). Наука (80-.). 357 , 1282 LP – 1287 (2017).
ADS Статья CAS Google Scholar
Брунер, Э., де ла Куэтара, Дж. М., Мастерс, М., Амано, Х. и Огихара, Н. Функциональная краниология и эволюция мозга: от палеонтологии к биомедицине. Фронт. Нейроанат. 8 , 19 (2014).
PubMed PubMed Central Статья Google Scholar
Бир, Дж. С., Джон, О.П., Скабини Д. и Найт Р. Т. Орбитофронтальная кора и социальное поведение: интеграция самоконтроля и эмоционально-познавательных взаимодействий. J. Cogn. Neurosci. 18 , 871–879 (2006).
PubMed Статья PubMed Central Google Scholar
Миллер, К. Дж., Ботвиник, М. М. и Броуди, К. Д. Ценностные представления в обучении орбитофронтальной коры головного мозга, но не выборе. bioRxiv (2018).
М. и Броуди, К. Д. Ценностные представления в обучении орбитофронтальной коры головного мозга, но не выборе. bioRxiv (2018).
Смит Б. Х. и Томпкинс Р. Л. К истории жизни гоминид. Annu. Преподобный Антрополь. 24 , 257–279 (1995).
Артикул Google Scholar
Бок, Дж. И Селлен, Д. У. Детство и эволюция жизненного пути человека. Hum. Nat. 13 , 153–159 (2002).
PubMed Статья PubMed Central Google Scholar
Смит, Т. М. и др. . Стоматологические доказательства онтогенетических различий между современными людьми и неандертальцами. Proc Natl Acad Sci USA 107 , 20923–20928 (2010).
ADS CAS PubMed Статья Google Scholar
Рамирес Роцци, Ф. В. и де Кастро, Дж. М. Удивительно быстрый рост неандертальцев. Природа 428 (2004).
Природа 428 (2004).
День, Ф.Р. и др. . Геномный анализ выявляет сотни вариантов, связанных с возрастом начала менархе, и подтверждает роль времени полового созревания в риске рака. Nat. Genet. 49 , 834–841 (2017).
CAS PubMed PubMed Central Статья Google Scholar
Antonson, P. et al. . Инактивация коактиватора ядерного рецептора RAP250 у мышей приводит к дисфункции плацентарных сосудов. Мол. Клетка. Биол. 23 , 1260–1268 (2003).
CAS PubMed PubMed Central Статья Google Scholar
Zhu, G.-Z., Lin, Y., Myles, D. G. & Primakoff, P. Идентификация четырех новых ADAM с потенциальной ролью в сперматогенезе и оплодотворении. Gene 234 , 227–237 (1999).
CAS PubMed Статья PubMed Central Google Scholar

Тан, С. и др. . Двуаллельные мутации в CFAP43 и CFAP44 вызывают мужское бесплодие с множественными морфологическими аномалиями жгутиков сперматозоидов. г. J. Hum. Genet. 100 , 854–864 (2017).
CAS PubMed PubMed Central Статья Google Scholar
Wang, Z., Widgren, E. E., Sivashanmugam, P., O’Rand, M. G., Richardson, R. T. Ассоциация эппина с семеногелином на сперматозоидах человека1. Biol. Репрод. 72 , 1064–1070 (2005).
CAS PubMed Статья PubMed Central Google Scholar
Эллерман, Д. А. и др. . Идзумо является частью мультибелкового семейства, члены которого образуют большие комплексы на сперматозоидах млекопитающих. Мол. Репрод. Dev. 76 , 1188–1199 (2009).
CAS PubMed PubMed Central Статья Google Scholar

Симонти, К. Н. и др. . Фенотипическое наследие смешения современных людей и неандертальцев. Наука (80-.). 351 , 737 LP – 741 (2016).
ADS Статья CAS Google Scholar
Даннеманн М., Прюфер К. и Келсо Дж. Функциональные последствия интрогрессии неандертальцев у современных людей. Genome Biol. 18 , 61 (2017).
PubMed PubMed Central Статья Google Scholar
Ши, Дж. и др. . Вариации последовательности гена мю-опиоидного рецептора (OPRM1), связанные с зависимостью человека от героина. Hum. Мутат. 19 , 459–460 (2002).
PubMed Статья CAS PubMed Central Google Scholar
Tan, E. et al. . Этническая принадлежность и вариант OPRM независимо предсказывают восприятие боли и контролируемое пациентом обезболивание при послеоперационной боли. Мол. Боль 5 , 32 (2009).
Мол. Боль 5 , 32 (2009).
PubMed PubMed Central Статья CAS Google Scholar
Пирс, Э., Влодарски, Р., Мачин, А. и Данбар, Р. И. М. Вариации генов рецепторов β-эндорфина, окситоцина и дофамина связаны с различными аспектами социальной жизни человека. Proc. Natl. Акад. Sci. 114 , 5300 LP – 5305 (2017).
Артикул CAS Google Scholar
Хабиб, А.М. и др. . Новое заболевание нечувствительности к боли у человека, вызванное точечной мутацией в ZFHX2. Мозг awx326-awx326 (2017).
Buiting, K. et al. . C15orf2 и новый некодирующий транскрипт из области синдрома Прадера-Вилли / Ангелмана обнаруживают моноаллельную экспрессию в мозге плода. Genomics 89 , 588–595 (2007).
CAS PubMed Статья PubMed Central Google Scholar

Макларен, W. и др. . Предиктор ансамблевого варианта эффекта. Genome Biol. 17 , 122 (2016).
PubMed PubMed Central Статья CAS Google Scholar
Кумар, П., Хеникофф, С. и Нг, П. С. Прогнозирование эффектов кодирования несинонимичных вариантов на функцию белка с использованием алгоритма SIFT. Nat. Protoc. 4 , 1073–1081 (2009).
CAS PubMed Статья PubMed Central Google Scholar
Аджубей И.А. и др. . Метод и сервер для предсказания повреждающих миссенс-мутаций. Nat. Методы 7 , 248–9 (2010).
CAS PubMed PubMed Central Статья Google Scholar
Кирхер, М. и др. . Общая схема оценки относительной патогенности генетических вариантов человека. Нат Генет 46 , 310–315 (2014).
CAS PubMed PubMed Central Статья Google Scholar
Ричи, Дж. Р. С., Данхэм, И., Зеггини, Э. и Фличек, П. Функциональная аннотация вариантов некодирующей последовательности. Nat. Методы 11 , 294–6 (2014).
CAS PubMed PubMed Central Статья Google Scholar
Lek, M. et al. . Анализ генетической изменчивости, кодирующей белок, у 60 706 человек. Природа 536 , 285–291 (2016).
CAS PubMed PubMed Central Статья Google Scholar
Патен, Б. и др. . Полногеномная реконструкция предков млекопитающих на нуклеотидном уровне. Genome Res. 18 , 1829–1843 (2008 г.).
CAS PubMed PubMed Central Статья Google Scholar

Ян, Г. и др. . Секвенирование генома и сравнение двух моделей животных-приматов, кроме яванского макак и китайских макак-резус. Nat. Biotechnol. 29 , 1019 (2011).
CAS PubMed Статья Google Scholar
Шерри, С. Т. и др. . dbSNP: база данных генетической изменчивости NCBI. Nucleic Acids Res. 29 , 308–311 (2001).
CAS PubMed PubMed Central Статья Google Scholar
Grantham, R. Формула разницы аминокислот для объяснения эволюции белков. Наука (80-.) . 185 (1974).
ADS CAS PubMed Статья Google Scholar
Ли, Х. Статистическая структура для вызова SNP, обнаружения мутаций, сопоставления ассоциаций и оценки генетических параметров популяции на основе данных секвенирования. Биоинформатика 27 , 2987–2993 (2011).
CAS PubMed PubMed Central Статья Google Scholar
Quinlan, A.R. & Hall, I.M.BEDTools: гибкий набор утилит для сравнения геномных характеристик. Биоинформатика 26 , 841–842 (2010).
CAS PubMed PubMed Central Статья Google Scholar
Huber, W. et al. . Организация высокопроизводительного геномного анализа с помощью Bioconductor. Нат. Мет 12 , 115–121 (2015).
CAS Статья Google Scholar
Лоуренс М., Джентльмен Р. и Кэри В. rtracklayer: пакет R для взаимодействия с браузерами генома. Биоинформатика 25 , 1841–1842 (2009).
CAS PubMed PubMed Central Статья Google Scholar
Durinck, S. et al. . BioMart и Bioconductor: мощная связь между биологическими базами данных и анализом данных микрочипов. Биоинформатика 21 , 3439–3440 (2005).
Биоинформатика 21 , 3439–3440 (2005).
CAS PubMed Статья PubMed Central Google Scholar
Zhang, H., Meltzer, P. & Davis, S. RCircos: пакет R для путевых участков Circos 2D. BMC Bioinformatics 14 , 244 (2013).
PubMed PubMed Central Статья Google Scholar
Синьорелл А. DescTools: Инструменты для описательной статистики.(2017).
Хан, С., Андрес, А. М., Маркес-Бонет, Т. и Кулвилм, М. Генетическая изменчивость у видов Pan определяется демографической историей и несет в себе функции, специфичные для клонов. Genome Biol. Evol . evz047, https://doi.org/10.1093/gbe/evz047 (2019).
PubMed PubMed Central Статья Google Scholar
Prüfer, K. et al. . FUNC: пакет для обнаружения значимых ассоциаций между наборами генов и онтологическими аннотациями. BMC Bioinformatics 8 , 41 (2007).
BMC Bioinformatics 8 , 41 (2007).
PubMed PubMed Central Статья CAS Google Scholar
Szklarczyk, D. et al. . База данных STRING в 2017 году: сети белок-белковых ассоциаций с контролируемым качеством, стали общедоступными. Nucleic Acids Res. 45 , D362 – D368 (2017).
CAS PubMed Статья PubMed Central Google Scholar
Эволюционные и медицинские последствия архаической интрогрессии в геномы современного человека
Гены (Базель).2018 июл; 9 (7): 358.
Population Genomics Group, Centre Nacional d’Anàlisi Genòmica, Centre de Regulació Genòmica (CRG-CNAG), Parc Científic de Barcelona, Baldiri Reixac 4, 08028 Барселона, Каталония, Испания
Получено в июне 2018 г. 15; Принято 11 июля 2018 г.
Лицензиат MDPI, Базель, Швейцария. Эта статья представляет собой статью в открытом доступе, распространяемую в соответствии с условиями лицензии Creative Commons Attribution (CC BY) (http://creativecommons. org/licenses/by/4.0/). Эта статья цитировалась другими статьями в PMC. .
org/licenses/by/4.0/). Эта статья цитировалась другими статьями в PMC. .Abstract
Демографическая история анатомически современного человека (AMH) включает в себя множественные миграционные события, вымирание популяций и генетические адаптации. По мере того, как полногеномные данные из полного секвенирования генома становятся все более многочисленными и доступными даже для вымерших гомининов, открываются новые идеи эволюционной истории нашего вида. В настоящее время известно, что AMH скрещивались с архаичными гомининами после того, как они покинули африканский континент. Геномы современного человека неафриканского происхождения содержат фрагменты архаичного происхождения.В этом обзоре основное внимание уделяется последствиям для приспособляемости архаичного скрещивания в современных популяциях людей. Мы обсуждаем новые идеи и проблемы, с которыми сталкиваются исследователи при интерпретации потенциального воздействия интрогрессии на приспособленность, и проверяем гипотезы о роли отбора в контексте здоровья и болезни.
Ключевые слова: архаическая интрогрессия, приспособленность, естественный отбор, неандерталец, денисовец, анатомически современные люди
1. Широкое скрещивание гомининов
Демографическая история анатомически современных людей (AMH) сложна и включает большое количество миграции, генетические примеси и интрогрессии, вымирание популяций и генетические адаптации, которые накладываются друг на друга как во времени, так и в пространстве (см.).Из-за этой сложности эволюционная история человечества все еще далека от полного понимания [1]. В течение последних 30 лет наиболее приемлемым демографическим сценарием для объяснения недавней эволюции AMH была модель Out of Africa (OOA). Согласно этой модели, АМГ возник в Африке около 100–200 тысяч лет назад (тыс. Лет назад) в Восточной Африке [2,3] и мигрировал в остальной мир примерно на 50–60 тыс. Лет назад [4,5,6,7]. Эта широко принятая датировка появления Homo sapiens была недавно оспорена Hublin et al. [8], которые обнаружили окаменелости AMH возрастом 300 тыс. Лет назад в Джебель-Ирхуде в Марокко [9]. Аналогичным образом, Herschkovitz et al. [10] описали челюстей H. sapiens от 177 до 194 тыс. Лет назад в пещере Мислия, Израиль. Все эти исследования предполагают, что члены клады H. sapiens покинули Африку раньше, чем предполагалось ранее, вероятно, в ходе нескольких волн миграции OOA на разных стадиях эволюции. Классический «чистый» OOA предполагает, что смешение с другими архаичными популяциями, такими как неандертальцы или денисовцы, присутствовавшие во время подъема AMH, либо не происходило, либо было незначительным [11].
[8], которые обнаружили окаменелости AMH возрастом 300 тыс. Лет назад в Джебель-Ирхуде в Марокко [9]. Аналогичным образом, Herschkovitz et al. [10] описали челюстей H. sapiens от 177 до 194 тыс. Лет назад в пещере Мислия, Израиль. Все эти исследования предполагают, что члены клады H. sapiens покинули Африку раньше, чем предполагалось ранее, вероятно, в ходе нескольких волн миграции OOA на разных стадиях эволюции. Классический «чистый» OOA предполагает, что смешение с другими архаичными популяциями, такими как неандертальцы или денисовцы, присутствовавшие во время подъема AMH, либо не происходило, либо было незначительным [11].
Генеалогическое древо четырех групп первых людей, живших в Евразии 50 000 лет назад, и предполагаемый поток генов между группами в результате скрещивания (на основе [12,13,14,15,16]). Показаны направление и предполагаемая величина предполагаемых событий потока генов. Длина ветвей и время генных потоков не масштабируются. Светло-фиолетовый цвет указывает на события интрогрессии от неизвестных архаичных популяций (Призраки).
Однако геномные исследования древней ДНК показали, что АМГ скрещивается с другими линиями гоминидов, такими как неандертальцы и денисовцы, присутствующие в Евразии с 300 до 30–50 тысячелетий назад.Примесь неандертальцев произошла около 37–86 тыс. Лет назад и, скорее всего, между 47 и 65 тыс. Лет назад [17,18,19,20]. Событие примешивания к денисову произошло примерно в такой же промежуток времени, ~ 44–54 тыс. Лет назад [21]. Полногеномные последовательности древних образцов [22] показали, что неафриканские популяции за пределами Океании несут от 1,8% до 2,6% ДНК неандертальцев () [13]. Как описано в [18], жители Восточной Азии несут несколько больше ДНК неандертальцев (от 2,3% до 2,6%), чем люди в Западной Евразии (от 1,8% до 2,4%).Напротив, ДНК, введенная в организм современного человека от денисовцев, обнаруживается в основном у австрало-меланезийцев, что может составлять до 6% денисовской ДНК в их геномах и, в меньшей степени, у жителей Южной Азии [23] и жителей Восточной Азии [24]. ]. Эти оценки являются средними по геному современного человека. Однако определенные области генома могут иметь степень неандертальского происхождения до 64% у европейцев и 62% у азиатов.
]. Эти оценки являются средними по геному современного человека. Однако определенные области генома могут иметь степень неандертальского происхождения до 64% у европейцев и 62% у азиатов.
Новые исследования, основанные на текущем генетическом разнообразии, предполагают, что события архаической интрогрессии в AMH действительно произошли после миграции из Африки с другими скрытыми «призрачными» архаическими популяциями [14].Основная трудность для вывода об архаической интрогрессии в африканских современных человеческих геномах из африканских архаических популяций в основном связана с отсутствием в настоящее время генетического материала от останков архаичных гомининов, который можно было бы использовать в качестве заместителя для изучения источника интрогрессии, как климат африканского континента не благоприятствует сохранению ДНК. Тем не менее, появляется все больше свидетельств того, что архаическая интрогрессия имела место и на этом континенте [15,25,26,27,28,29], что поднимает захватывающую возможность того, что другие неизвестные архаические группы могли внести свой вклад в генетическое разнообразие человека. Таким образом, недавняя работа предполагает, что явно отдельные виды могут обмениваться генетическим материалом на протяжении своей эволюционной истории [30]. Биологические последствия такой интрогрессии, включая их последствия для здоровья современного человека, рассматриваются в следующих разделах.
Таким образом, недавняя работа предполагает, что явно отдельные виды могут обмениваться генетическим материалом на протяжении своей эволюционной истории [30]. Биологические последствия такой интрогрессии, включая их последствия для здоровья современного человека, рассматриваются в следующих разделах.
2. Селекция против интрогрессированных областей на уровне геномов и индивидуальных локусов
Интрогрессированные аллели в чужеродном генетическом фоне часто имеют отрицательные эффекты приспособленности независимо от степени адаптивной интрогрессии.Мартин и Джиггинс [31] сделали два важных соображения, имея дело с моделями отбора против интрогрессированных геномных трактов. Во-первых, поскольку многие факторы, влияющие на отбор, такие как скорость рекомбинации и плотность генов, являются взаимозависимыми, модели, которые учитывают комбинированные эффекты обоих факторов, более осуществимы, особенно если они включают конкретные прогнозы, такие как снижение силы выборочной развертки. по мере удаления от выбранных локусов. Юрич и др. [32] смоделировали уровень неандертальского происхождения в человеческих популяциях как функцию расстояния рекомбинации от ближайших выбранных аллелей и оценили как плотность выбранных локусов, так и силу отбора.Во-вторых, интерпретация пейзажа древних вторжений в человеческий вид может варьироваться в зависимости от сделанных допущений. Например, большинство моделей предполагают, что интрогрессированные блоки выбираются независимо друг от друга в генетическом фоне популяции-реципиента. Однако Харрис и Нильсен [33] показали, что большая часть отбора против интрогрессии может происходить в ранних поколениях, поскольку гибриды ранних поколений должны иметь сложные предки, в которых эпистаз может приводить к неаддитивным эффектам приспособленности.Другое предположение, указанное в этом исследовании, заключается в том, что слабо вредные мутации, сегрегация в популяции доноров, будут основным фактором отбора против интрогрессии неандертальцев у людей.
по мере удаления от выбранных локусов. Юрич и др. [32] смоделировали уровень неандертальского происхождения в человеческих популяциях как функцию расстояния рекомбинации от ближайших выбранных аллелей и оценили как плотность выбранных локусов, так и силу отбора.Во-вторых, интерпретация пейзажа древних вторжений в человеческий вид может варьироваться в зависимости от сделанных допущений. Например, большинство моделей предполагают, что интрогрессированные блоки выбираются независимо друг от друга в генетическом фоне популяции-реципиента. Однако Харрис и Нильсен [33] показали, что большая часть отбора против интрогрессии может происходить в ранних поколениях, поскольку гибриды ранних поколений должны иметь сложные предки, в которых эпистаз может приводить к неаддитивным эффектам приспособленности.Другое предположение, указанное в этом исследовании, заключается в том, что слабо вредные мутации, сегрегация в популяции доноров, будут основным фактором отбора против интрогрессии неандертальцев у людей. При такой модели более низкий эффективный размер популяции неандертальцев привел бы к накоплению слабо вредных аллелей, которые, будучи внедренными в организм человека, снизили бы относительную приспособленность гибрида. Однако в таком контексте, даже если оба вида несут рецессивные вредные аллели, но на разных участках, гибриды могли усилить сверхдоминантную изменчивость приспособленности независимо от этих вредных рецессивов, что приводит к выводу, что интрогрессия неандертальцев, возможно, изначально благоприятствовала отбору у людей [33 ].
При такой модели более низкий эффективный размер популяции неандертальцев привел бы к накоплению слабо вредных аллелей, которые, будучи внедренными в организм человека, снизили бы относительную приспособленность гибрида. Однако в таком контексте, даже если оба вида несут рецессивные вредные аллели, но на разных участках, гибриды могли усилить сверхдоминантную изменчивость приспособленности независимо от этих вредных рецессивов, что приводит к выводу, что интрогрессия неандертальцев, возможно, изначально благоприятствовала отбору у людей [33 ].
Непропорциональная роль половых хромосом в видовом различии и несовместимости гибридов составляет постоянный образец видообразования [34,35]. Неопровержимые доказательства этих процессов были получены в геномах людей неафриканского происхождения, которые имеют последовательности, лишенные интрогрессивной вариации («пустыни») от неандертальцев и денисовцев, возможно, вызванные отбором против интрогрессии, описанным Sankararaman et al. [21,36]. Более того, авторы указали, что пустыни интрогрессии ДНК неандертальцев и денисовцев у современных людей в значительной степени перекрываются.Особый интерес представляет значительное уменьшение примеси, связанной с генами, показывающими специфическую для семенников экспрессию, предполагая, что примесь могла привести к снижению мужской фертильности, и подтверждая доказательства снижения интрогрессии половых хромосом [21,36,37]. Однако это геномное свидетельство следует интерпретировать с осторожностью [38]. Когда отбор против интрогрессии происходит в большом количестве локусов по всему геному, его комбинированное воздействие на многие локусы может оставлять обнаруживаемые паттерны, даже если отбор по любому отдельному локусу может быть слабым [31].Более того, более слабые сигналы интрогрессии наблюдались в частях генома с высокой плотностью генов и / или низкой рекомбинацией [21], что согласуется с теоретической работой, которая предсказывала, что сила отбора против интрогрессии зависит от плотности выбранных сайтов и скорость рекомбинации [39].
Более того, авторы указали, что пустыни интрогрессии ДНК неандертальцев и денисовцев у современных людей в значительной степени перекрываются.Особый интерес представляет значительное уменьшение примеси, связанной с генами, показывающими специфическую для семенников экспрессию, предполагая, что примесь могла привести к снижению мужской фертильности, и подтверждая доказательства снижения интрогрессии половых хромосом [21,36,37]. Однако это геномное свидетельство следует интерпретировать с осторожностью [38]. Когда отбор против интрогрессии происходит в большом количестве локусов по всему геному, его комбинированное воздействие на многие локусы может оставлять обнаруживаемые паттерны, даже если отбор по любому отдельному локусу может быть слабым [31].Более того, более слабые сигналы интрогрессии наблюдались в частях генома с высокой плотностью генов и / или низкой рекомбинацией [21], что согласуется с теоретической работой, которая предсказывала, что сила отбора против интрогрессии зависит от плотности выбранных сайтов и скорость рекомбинации [39].
Доказательства роли очищающего отбора в формировании ландшафта интрогрессии исходят из конкретных категорий генов, испытывающих различную степень интрогрессии, как ранее было продемонстрировано для нечеловеческих видов [40,41,42,43].Это также верно для аутосомных регионов, дефицитных как у неандертальцев, так и у денисовских предков, которые содержат значительное обогащение генов, транскрибируемых в мейотических зародышевых клетках [44,45]. Фенотипические признаки, на которые влияет архаическая интрогрессия, обобщены, а соответствующие им участки генома с типом действующего на них режима отбора перечислены в Таблице S1. Принимая во внимание тот факт, что между генами, кодирующими белок [21,46,47], был проведен строгий отбор против архаической интрогрессии, функциональные области, вносящие вклад в уникальность некоторых черт современного человека, могут быть идентифицированы, если они сильно лишены архаической родословной [48]. .Например, не было обнаружено ни одного предка неандертальцев вокруг гена белка P2 ( FOXP2 ) [21], мутации которого связаны с языковыми расстройствами [49]. Точно так же неандертальцы и денисовцы несут единственную копию гена AMY1 , кодирующего фермент амилазу, ответственный за переваривание крахмала [21]. Напротив, AMH несут множественные копии гена [50,51], и нет никаких доказательств интрогрессии неандертальцев [21]. Это было интерпретировано как доказательство того, что производство большего количества амилазы слюны для переваривания крахмала у современных людей подвергалось положительному отбору по сравнению с архаическими видами [52].Более того, области, обедненные как предками неандертальцев, так и денисовых, обогащены генами, экспрессируемыми в определенных областях мозга (например, вентральная лобная кора — вентролатеральная префронтальная кора у младенцев и полосатое тело во взрослом возрасте; [48]). Другое геномное исследование Chintalapati et al. [53] на небольших инделах, интрогрессированных от неандертальца, продемонстрировали, что отрицательный отбор влияет на эти варианты больше, чем на другие сегрегационные варианты у современных людей, и подтвердил, что делеции развивались с большим ограничением, чем вставки, подавляющее большинство из которых лежало в интронных регионах.
Точно так же неандертальцы и денисовцы несут единственную копию гена AMY1 , кодирующего фермент амилазу, ответственный за переваривание крахмала [21]. Напротив, AMH несут множественные копии гена [50,51], и нет никаких доказательств интрогрессии неандертальцев [21]. Это было интерпретировано как доказательство того, что производство большего количества амилазы слюны для переваривания крахмала у современных людей подвергалось положительному отбору по сравнению с архаическими видами [52].Более того, области, обедненные как предками неандертальцев, так и денисовых, обогащены генами, экспрессируемыми в определенных областях мозга (например, вентральная лобная кора — вентролатеральная префронтальная кора у младенцев и полосатое тело во взрослом возрасте; [48]). Другое геномное исследование Chintalapati et al. [53] на небольших инделах, интрогрессированных от неандертальца, продемонстрировали, что отрицательный отбор влияет на эти варианты больше, чем на другие сегрегационные варианты у современных людей, и подтвердил, что делеции развивались с большим ограничением, чем вставки, подавляющее большинство из которых лежало в интронных регионах. Кроме того, были выявлены интрогрессированные варианты, которые могут влиять на фенотип их носителей (таблица S1). Среди них интрогрессированная делеция, связанная с уменьшением времени до менархе, может представлять собой пример бывшей неандертальской особенности, вносящей вклад в фенотипическое разнообразие современного человека [53] (Таблица S1).
Кроме того, были выявлены интрогрессированные варианты, которые могут влиять на фенотип их носителей (таблица S1). Среди них интрогрессированная делеция, связанная с уменьшением времени до менархе, может представлять собой пример бывшей неандертальской особенности, вносящей вклад в фенотипическое разнообразие современного человека [53] (Таблица S1).
Современные человеческие органы и системы, пораженные интрогрессированными вариантами из древних геномов (подробности см. В Таблице S1).
Дальнейшие доказательства пагубного эффекта интрогрессии неандертальцев могут быть идентифицированы на уровне экспрессии.Анализ экспрессии генов аллелей неандертальцев у нынешних людей показывает значительное подавление в яичках и головном мозге по сравнению с другими тканями [54,55].
3. Геномные сигнатуры адаптивной интрогрессии от архаичного к современному человеку
След очищающего отбора против архаичных аллелей широко распространен в геноме человека. Тем не менее, учитывая, что архаические виды эволюционировали в течение долгого времени в средах, для которых ранний AMH не был биологически адаптирован, скрещивание анатомически современных людей с архаичными видами могло облегчить адаптацию к специфическим средам [56,57] (см. Таблицу S1).Этот эволюционный процесс может приводить варианты с большей частотой, чем мутации de novo, обеспечивая связанные блоки последовательности с множественными функциональными мутациями, потенциально включая коадаптированные аллели [58]. Этот процесс, известный как адаптивная интрогрессия, приобрел известность на основе ряда ярких примеров в геномах человека [56,59,60] (см.). Например, гены, участвующие в функциях, связанных с кератиновыми нитями, метаболизмом сахара, сокращением мышц, распределением жировых отложений, толщиной эмали, мейозом ооцитов, размером мозга и функционированием, стали мишенью адаптивной интрогрессии неандертальцев в различных неафриканских геномах [29,36 , 56,61,62,63,64,65].Гены, участвующие в изменении пигментации кожи и морфологии волос ( BNC2, MC1R ), также демонстрируют признаки положительного отбора как результат адаптации к различным средам обитания с разной степенью инсоляции (Таблица S1) [54,66,67]. Полезные иммунные варианты, введенные в современную человеческую популяцию из архаичных геномов, внесли значительный вклад в современное разнообразие иммунных генов [56,68,69,70,71,72].
Таблицу S1).Этот эволюционный процесс может приводить варианты с большей частотой, чем мутации de novo, обеспечивая связанные блоки последовательности с множественными функциональными мутациями, потенциально включая коадаптированные аллели [58]. Этот процесс, известный как адаптивная интрогрессия, приобрел известность на основе ряда ярких примеров в геномах человека [56,59,60] (см.). Например, гены, участвующие в функциях, связанных с кератиновыми нитями, метаболизмом сахара, сокращением мышц, распределением жировых отложений, толщиной эмали, мейозом ооцитов, размером мозга и функционированием, стали мишенью адаптивной интрогрессии неандертальцев в различных неафриканских геномах [29,36 , 56,61,62,63,64,65].Гены, участвующие в изменении пигментации кожи и морфологии волос ( BNC2, MC1R ), также демонстрируют признаки положительного отбора как результат адаптации к различным средам обитания с разной степенью инсоляции (Таблица S1) [54,66,67]. Полезные иммунные варианты, введенные в современную человеческую популяцию из архаичных геномов, внесли значительный вклад в современное разнообразие иммунных генов [56,68,69,70,71,72]. Поскольку гены врожденного иммунитета эволюционировали в условиях более сильного очищающего отбора, чем остальная часть генома [73], такое обогащение интрогрессированных аллелей предполагает наличие сильного положительного отбора в иммунной системе.Более широкий обзор древних патогенов, передающихся в геномы современного человека через половые взаимодействия с архаичными гомининами, и их влияние на эволюцию иммунитета к AMH можно найти в обзоре Pimenoff et al. [74] опубликованы в этом специальном выпуске по эволюционной медицине. Подобно генам врожденного иммунитета, гены EGLN1 и EPAS1 , связанные с концентрацией гемоглобина и реакцией на гипоксию, демонстрируют высокую степень денисовского происхождения у тибетцев, что позволяет предположить, что эта популяция приобрела выгодные аллели для жизни на большой высоте за счет древней примеси [75 , 76,77].В отличие от этих доказательств положительного отбора, доказательства сбалансированного отбора у людей в основном косвенные [78].
Поскольку гены врожденного иммунитета эволюционировали в условиях более сильного очищающего отбора, чем остальная часть генома [73], такое обогащение интрогрессированных аллелей предполагает наличие сильного положительного отбора в иммунной системе.Более широкий обзор древних патогенов, передающихся в геномы современного человека через половые взаимодействия с архаичными гомининами, и их влияние на эволюцию иммунитета к AMH можно найти в обзоре Pimenoff et al. [74] опубликованы в этом специальном выпуске по эволюционной медицине. Подобно генам врожденного иммунитета, гены EGLN1 и EPAS1 , связанные с концентрацией гемоглобина и реакцией на гипоксию, демонстрируют высокую степень денисовского происхождения у тибетцев, что позволяет предположить, что эта популяция приобрела выгодные аллели для жизни на большой высоте за счет древней примеси [75 , 76,77].В отличие от этих доказательств положительного отбора, доказательства сбалансированного отбора у людей в основном косвенные [78]. Однако гены защиты хозяина, такие как гены, кодирующие несколько мембранных гликопротеинов, области KIR , которые совместно эволюционируют с лигандами HLA , и другие гены, кодирующие белки, участвующие в миграции клеток или врожденном иммунитете, по-видимому, подвергаются этому редкому в других отношениях селективному режиму [79 , 80,81,82,83]. Область HLA , парадигма балансирующего отбора у людей, содержит функциональные варианты, которые, вероятно, были интрогрессированы от неандертальцев и денисовцев [84].Альтернативное объяснение Ясукочи и Охаши [85], основанное на филогенетическом анализе, не поддерживает гипотезу интрогрессии и заключает, что весьма вероятно, что предположительно интрогрессированная аллельная линия HLA-B * 73 сохранилась у прямых предков современных людей. [85].
Однако гены защиты хозяина, такие как гены, кодирующие несколько мембранных гликопротеинов, области KIR , которые совместно эволюционируют с лигандами HLA , и другие гены, кодирующие белки, участвующие в миграции клеток или врожденном иммунитете, по-видимому, подвергаются этому редкому в других отношениях селективному режиму [79 , 80,81,82,83]. Область HLA , парадигма балансирующего отбора у людей, содержит функциональные варианты, которые, вероятно, были интрогрессированы от неандертальцев и денисовцев [84].Альтернативное объяснение Ясукочи и Охаши [85], основанное на филогенетическом анализе, не поддерживает гипотезу интрогрессии и заключает, что весьма вероятно, что предположительно интрогрессированная аллельная линия HLA-B * 73 сохранилась у прямых предков современных людей. [85].
Все больше данных свидетельствует о том, что регуляторные варианты играют центральную роль в адаптивных процессах [86,87,88]. Убедительным примером локальной адаптации, обнаруженной на уровне экспрессии, является ген рецептора апелина APLNR . Апелин — это сигнальный пептид, который влияет на несколько аспектов сердечной, пищеварительной, мозговой и сосудистой функций, включая регуляцию уровня кислорода. Этот ген демонстрирует сильную аллель-специфическую экспрессию, благоприятствующую аллелю неандертальца в тканях мозга, но аллель-специфичную экспрессию, благоприятствующую аллелю современного человека в тканях, не относящихся к мозгу [55]. Существует также ряд примеров местной адаптации, вызванной регуляторными вариантами, приводящими к популяционным различиям в иммунных ответах [71,72,89]. Несмотря на доказательства того, что функциональные архаичные аллели (несинонимичные и связанные с экспрессией) снизились по частоте за последние 8500 лет, были идентифицированы четыре локуса, в которых архаичные аллели, связанные с дифференциальной экспрессией, показывают значительное увеличение частоты с течением времени.Среди них интрогрессированные аллели, модифицирующие экспрессию генов OAS1 / OAS2 / OAS3 , участвующих в врожденном иммунитете и чьи тканеспецифические эффекты предполагают, что они могут иметь функциональное значение [71,89].
Апелин — это сигнальный пептид, который влияет на несколько аспектов сердечной, пищеварительной, мозговой и сосудистой функций, включая регуляцию уровня кислорода. Этот ген демонстрирует сильную аллель-специфическую экспрессию, благоприятствующую аллелю неандертальца в тканях мозга, но аллель-специфичную экспрессию, благоприятствующую аллелю современного человека в тканях, не относящихся к мозгу [55]. Существует также ряд примеров местной адаптации, вызванной регуляторными вариантами, приводящими к популяционным различиям в иммунных ответах [71,72,89]. Несмотря на доказательства того, что функциональные архаичные аллели (несинонимичные и связанные с экспрессией) снизились по частоте за последние 8500 лет, были идентифицированы четыре локуса, в которых архаичные аллели, связанные с дифференциальной экспрессией, показывают значительное увеличение частоты с течением времени.Среди них интрогрессированные аллели, модифицирующие экспрессию генов OAS1 / OAS2 / OAS3 , участвующих в врожденном иммунитете и чьи тканеспецифические эффекты предполагают, что они могут иметь функциональное значение [71,89]. Архаичные аллели OAS1 связаны с более высокой экспрессией в подкожной жировой ткани и на коже, подвергающейся воздействию солнца, в то время как более высокая экспрессия OAS2 в щитовидной железе и OAS3 в поджелудочной железе и влагалище связана с архаичными аллелями.Напротив, у лиц, несущих архаические аллели, наблюдается подавление регуляции OAS1 в слизистой оболочке пищевода и селезенки, OAS2 в фибробластах и OAS3 в фибробластах, а также в слизистой оболочке пищевода, селезенке и трех областях мозга (гиппокампе, гиппокампе, и хвостатое ядро) [84]. Другие примеры местной адаптации, влияющей на уровни экспрессии, включают экспрессию гена ERAP2 , вовлеченного в предрасположенность к болезни Крона; CCR1 , ограничивающий рекрутирование лейкоцитов и предотвращающий воспалительные реакции; HLA-DQA1 , связанный с предрасположенностью к целиакии; и TLR1 , связанный с заметно более низкими уровнями экспрессии гена воспалительного ответа [71,72].
Архаичные аллели OAS1 связаны с более высокой экспрессией в подкожной жировой ткани и на коже, подвергающейся воздействию солнца, в то время как более высокая экспрессия OAS2 в щитовидной железе и OAS3 в поджелудочной железе и влагалище связана с архаичными аллелями.Напротив, у лиц, несущих архаические аллели, наблюдается подавление регуляции OAS1 в слизистой оболочке пищевода и селезенки, OAS2 в фибробластах и OAS3 в фибробластах, а также в слизистой оболочке пищевода, селезенке и трех областях мозга (гиппокампе, гиппокампе, и хвостатое ядро) [84]. Другие примеры местной адаптации, влияющей на уровни экспрессии, включают экспрессию гена ERAP2 , вовлеченного в предрасположенность к болезни Крона; CCR1 , ограничивающий рекрутирование лейкоцитов и предотвращающий воспалительные реакции; HLA-DQA1 , связанный с предрасположенностью к целиакии; и TLR1 , связанный с заметно более низкими уровнями экспрессии гена воспалительного ответа [71,72]. По-видимому, интрогрессия со стороны неандертальцев также способствовала диверсификации транскрипционных ответов на инфекцию в человеческих популяциях. Интрогрессированные генетические сегменты в европейских геномах содержат регуляторные варианты с эффектами на устойчивую экспрессию и ответы на стимуляцию TLR7 / 8 и вирус гриппа [72,78]. Кроме того, об архаичных вариантах нескольких локусов количественных признаков экспрессии (eQTL) сообщалось как о потенциальных кандидатах на адаптивную интрогрессию, обеспечивающую лучшую адаптацию за счет регуляции экспрессии генов.Примерами являются ген DARS , связанный с нейровоспалительными расстройствами и расстройствами белого вещества [71]; архаичные варианты локуса OAS , по-видимому, ассоциированы с различными фенотипами устойчивости к флавивирусам [90]; и PNMA1 , несущий eQTL ответа на вирус гриппа и стимулирующий продукцию интерферона [72]. Другой регуляторный архаичный вариант, который модифицирует экспрессию TNPO3 в головном мозге, связан с множественными аутоиммунными фенотипами [83].
По-видимому, интрогрессия со стороны неандертальцев также способствовала диверсификации транскрипционных ответов на инфекцию в человеческих популяциях. Интрогрессированные генетические сегменты в европейских геномах содержат регуляторные варианты с эффектами на устойчивую экспрессию и ответы на стимуляцию TLR7 / 8 и вирус гриппа [72,78]. Кроме того, об архаичных вариантах нескольких локусов количественных признаков экспрессии (eQTL) сообщалось как о потенциальных кандидатах на адаптивную интрогрессию, обеспечивающую лучшую адаптацию за счет регуляции экспрессии генов.Примерами являются ген DARS , связанный с нейровоспалительными расстройствами и расстройствами белого вещества [71]; архаичные варианты локуса OAS , по-видимому, ассоциированы с различными фенотипами устойчивости к флавивирусам [90]; и PNMA1 , несущий eQTL ответа на вирус гриппа и стимулирующий продукцию интерферона [72]. Другой регуляторный архаичный вариант, который модифицирует экспрессию TNPO3 в головном мозге, связан с множественными аутоиммунными фенотипами [83]. Все эти исследования ясно показывают, что отбор и архаичное смешение существенно влияют на современные межпопуляционные различия в иммунных ответах, по крайней мере, с точки зрения транскрипционной изменчивости, подтверждая представление о том, что вариации в экспрессии генов были важным средством адаптации человека [86]. .Более того, было показано, что архаические варианты с более высокой частотой вносят значительно больший вклад в изменения экспрессии генов, чем архаические варианты с более низкой частотой. Это предполагает, что по крайней мере некоторые из архаичных аллелей, которые модифицируют экспрессию генов, могли быть доведены до более высоких частот во многих человеческих популяциях благодаря положительному отбору, подтверждая идею о том, что изменения в экспрессии генов, вероятно, будут иметь важные адаптивные эффекты у людей [89].
Все эти исследования ясно показывают, что отбор и архаичное смешение существенно влияют на современные межпопуляционные различия в иммунных ответах, по крайней мере, с точки зрения транскрипционной изменчивости, подтверждая представление о том, что вариации в экспрессии генов были важным средством адаптации человека [86]. .Более того, было показано, что архаические варианты с более высокой частотой вносят значительно больший вклад в изменения экспрессии генов, чем архаические варианты с более низкой частотой. Это предполагает, что по крайней мере некоторые из архаичных аллелей, которые модифицируют экспрессию генов, могли быть доведены до более высоких частот во многих человеческих популяциях благодаря положительному отбору, подтверждая идею о том, что изменения в экспрессии генов, вероятно, будут иметь важные адаптивные эффекты у людей [89].
Однако, какими бы ни были потенциальные преимущества архаической интрогрессии в прошлом, аллели неандертальского происхождения также были связаны с несколькими неврологическими, дерматологическими и иммунологическими фенотипами, что указывает на влияние древних примесей на текущий риск заболеваний у людей [91,92, 93]. Например, интрогрессированные аллели, связанные с ответом иммунной системы, могут увеличивать риск воспаления или аутоиммунитета при изменении факторов окружающей среды со временем [94,95,96,97,98]. Случай целиакии наглядно иллюстрирует компромисс между прошлым отбором и текущей дезадаптацией. Taskent et al. [93] обнаружили свидетельства интрогрессии неандертальцев в семейство генов хемокиновых рецепторов ( CCR ), составляющих аллели риска глютеновой болезни, что, возможно, поддерживалось селективными силами в ранней европейской популяции.Кроме того, популяционный генетический анализ показал, что высокая частота нескольких аллелей риска генов, связанных с глютеновой болезнью, таких как IL12A , IL18RAP и Sh3B3 [99], у европейцев является результатом прошлых событий положительного отбора [94,100]. Другой пример — несинонимичный вариант гена ZNF365D , присутствующий у ~ 32% европейцев и отсутствующий у африканцев, унаследованный от неандертальцев и связанный с более высоким риском болезни Крона [36].
Например, интрогрессированные аллели, связанные с ответом иммунной системы, могут увеличивать риск воспаления или аутоиммунитета при изменении факторов окружающей среды со временем [94,95,96,97,98]. Случай целиакии наглядно иллюстрирует компромисс между прошлым отбором и текущей дезадаптацией. Taskent et al. [93] обнаружили свидетельства интрогрессии неандертальцев в семейство генов хемокиновых рецепторов ( CCR ), составляющих аллели риска глютеновой болезни, что, возможно, поддерживалось селективными силами в ранней европейской популяции.Кроме того, популяционный генетический анализ показал, что высокая частота нескольких аллелей риска генов, связанных с глютеновой болезнью, таких как IL12A , IL18RAP и Sh3B3 [99], у европейцев является результатом прошлых событий положительного отбора [94,100]. Другой пример — несинонимичный вариант гена ZNF365D , присутствующий у ~ 32% европейцев и отсутствующий у африканцев, унаследованный от неандертальцев и связанный с более высоким риском болезни Крона [36]. Точно так же варианты кластера генов TLR6-1-10 , унаследованные от неандертальцев и денисовцев и присутствующие у европейцев и азиатов, были связаны с большей предрасположенностью к аллергии [101].
Точно так же варианты кластера генов TLR6-1-10 , унаследованные от неандертальцев и денисовцев и присутствующие у европейцев и азиатов, были связаны с большей предрасположенностью к аллергии [101].
Требуются дальнейшие исследования, но исследования, опубликованные на сегодняшний день, предоставили неоценимые ресурсы и расширили наше понимание молекулярных и клеточных процессов, определяемых интрогрессированными генетическими вариантами и различными режимами отбора, действующими на них.
4.Выводы и перспективы
Продолжающийся поток данных секвенирования от тысяч людей и различных популяций по всему миру, включая некоторые архаичные гоминины и древние геномы AMH, предоставил новое понимание эволюционной истории нашего вида. Геномные исследования интрогрессии между ранними евразийцами и архаическими человеческими видами, такими как неандертальцы и денисовцы, начинают предлагать новое понимание эволюционных и фенотипических последствий гибридизации.Существует довольно распространенное свидетельство широко распространенного отбора против интрогрессии по всему геному, но адаптивная интрогрессия также может считаться важной движущей силой адаптации современных людей к новым условиям. Однако развитие дополнительных наборов человеческих данных, интеграция различных источников информации и разработка новых статистических и аналитических методов имеют решающее значение для понимания биологических и медицинских последствий таких сигналов выбора.
Однако развитие дополнительных наборов человеческих данных, интеграция различных источников информации и разработка новых статистических и аналитических методов имеют решающее значение для понимания биологических и медицинских последствий таких сигналов выбора.
Таблица S1 показывает, что наши знания о функциональных последствиях интрогрессивной вариации в основном основаны на популяциях европейского происхождения.Информативные данные из других этнических групп и секвенирование дополнительных образцов от древних гомининов еще больше углубят наши знания о вкладе архаичных гомининов в разнообразие человеческих черт и сложных заболеваний. Кроме того, это поможет выявить функциональные изменения, которые с течением времени способствовали адаптации и выживанию человека. Более того, перспективные когортные исследования нескольких поколений из множества человеческих популяций позволят проводить прямые измерения генетической изменчивости и интенсивности отбора общих черт в современных популяциях, проводимые в различных пищевых, культурных и географических условиях, что представляет собой лучший способ охарактеризовать масштабы и важность сложных экологических, эпидемиологических, демографических и эволюционных сдвигов. В дополнение к базам данных геномов европейского происхождения, таких как когорта Framingham в США, когорта рождения Uppsala в Швеции и когорта Lifelines в Нидерландах, такие когорты теперь включают инициативу h4Africa по наследственности и здоровью человека в Африке и проект Tohoku Medical Megabank. в Японии. Необходимые данные, дополненные геномной и медицинской информацией, будут способствовать дальнейшему углублению нашего понимания антагонистических плейотропных эффектов, которые вносят вклад в бремя неинфекционных заболеваний, и предоставят новые ключи к разгадке причин заболевания, потенциальным методам лечения и возможным побочным эффектам новых методов лечения [96].
В дополнение к базам данных геномов европейского происхождения, таких как когорта Framingham в США, когорта рождения Uppsala в Швеции и когорта Lifelines в Нидерландах, такие когорты теперь включают инициативу h4Africa по наследственности и здоровью человека в Африке и проект Tohoku Medical Megabank. в Японии. Необходимые данные, дополненные геномной и медицинской информацией, будут способствовать дальнейшему углублению нашего понимания антагонистических плейотропных эффектов, которые вносят вклад в бремя неинфекционных заболеваний, и предоставят новые ключи к разгадке причин заболевания, потенциальным методам лечения и возможным побочным эффектам новых методов лечения [96].
В случае интеграции различных источников информации, исследования генетических вариантов с регуляторными эффектами на экспрессию генов (eQTL) уже предоставили понимание генетических и эволюционных детерминант фенотипического разнообразия популяций [102]. Одним из первых глобальных подходов является проект Genotype-Tissue Expression (GTEx). GTEx исследует ландшафт экспрессии генов в 54 различных тканях, обеспечивая богатейший каталог тканеспецифичных и общих eQTL [103,104].Он уже использовался при анализе паттернов экспрессии интрогрессированных гаплотипов в недавних исследованиях, проведенных McCoy et al. [55] и Dannemann et al. [89]. Будущий популяционно-генетический анализ должен распространить это повсеместное обоснование на несколько популяций разного этнического происхождения, чтобы получить полную картину физиологических механизмов, лежащих в основе адаптации к давлению окружающей среды и поддержания гомеостаза.
GTEx исследует ландшафт экспрессии генов в 54 различных тканях, обеспечивая богатейший каталог тканеспецифичных и общих eQTL [103,104].Он уже использовался при анализе паттернов экспрессии интрогрессированных гаплотипов в недавних исследованиях, проведенных McCoy et al. [55] и Dannemann et al. [89]. Будущий популяционно-генетический анализ должен распространить это повсеместное обоснование на несколько популяций разного этнического происхождения, чтобы получить полную картину физиологических механизмов, лежащих в основе адаптации к давлению окружающей среды и поддержания гомеостаза.
В случае новых методов задача на будущее будет заключаться в разработке надежных статистических моделей и вычислительных методов для обнаружения отбора, более широкого количественного определения частоты адаптивной интрогрессии и понимания обстоятельств, при которых она, вероятно, будет играть преобладающую роль в приспособление.Захватывающая перспектива будущего состоит в том, что нашей интерпретации наблюдений в природе будут помогать имитационные исследования [33] и эмпирические исследования последствий интрогрессии для фенотипа и приспособленности [55]. В этом контексте нет точных оценок времени появления большинства признаков отбора, обнаруживаемых в настоящее время в геноме человека (персистентность лактазы в качестве исключения) [105,106], или хороших методов для оценки возраста и природы окружающей среды в который произошел в прошлом отборе [97].Будущие исследования должны иметь возможность с уверенностью различать разные временные шкалы отбора, например, в результате сельскохозяйственной революции 8000–10 000 лет назад или промышленной революции 100–300 лет назад.
В заключение, интеграция всех этих наборов данных в структуру клинической, эпидемиологической и популяционной генетики позволит по-новому взглянуть на историю адаптации рода Homo , а также на то, как наш генетический и негенетический состав вместе взятые. Изменения в окружающей среде и культурных традициях влияют на фенотипические различия как в состоянии здоровья, так и в отношении болезней.
Благодарности
Мы благодарим Антонио Барбадиллу и двух анонимных рецензентов за полезные комментарии к рукописи.
Вклад авторов
O.D. отобрана соответствующая литература для обзора; О. и О. просмотрел литературу и написал рукопись.
Финансирование
Эта работа была поддержана грантом BFU2015-68759-P (MINECO / FEDER) и при поддержке Secretaria d’Universitats i Recerca del Departament d’Economia i Coneixement de la Generalitat de Catalunya (GRC 2017 SGR 937) .O.L также была поддержана грантом Рамона и Кахала от Министерства экономики и конкуренции Испании (MINECO) со ссылкой RYC-2013-14797.
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов.
Ссылки
1. Нильсен Р., Эйки Дж. М., Якобссон М., Притчард Дж. К., Тишкофф С., Виллерслев Э. Прослеживание населения мира с помощью геномики. Природа. 2017; 541: 302–310. DOI: 10,1038 / природа21347. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 2.Уайт Т.Д., Асфау Б., ДеГуста Д., Гилберт Х., Ричардс Г.Д., Сува Г., Хауэлл F.C. Плейстоцен Homo sapiens из Среднего Аваша, Эфиопия. Природа. 2003; 423: 742–747. DOI: 10,1038 / природа01669. [PubMed] [CrossRef] [Google Scholar] 3. Макдугалл И., Браун Ф. Х., Флигл Дж. Стратиграфическое расположение и возраст современных людей из Кибиша, Эфиопия. Природа. 2005; 433: 733–736. DOI: 10,1038 / природа03258. [PubMed] [CrossRef] [Google Scholar] 4. Мерсье Н., Валладас Х., Бар-Йосеф О., Вандермерш Б., Стрингер К., Джорон Ж.-Л. Дата термолюминесценции мустьерского могильника Эс-Схул, гора Кармель. J. Archaeol. Sci. 1993. 20: 169–174. DOI: 10.1006 / jasc.1993.1012. [CrossRef] [Google Scholar] 5. Стрингер К. Природа. 1989; 338: 756–758. DOI: 10.1038 / 338756a0. [PubMed] [CrossRef] [Google Scholar] 6. Schwarcz H.P., Grün R., Vandermeersch B., Bar-Yosef O., Valladas H., Tchernov E. Даты ЭПР для захоронения гоминидов в Кафзе в Израиле.Гм. Evol. 1988; 17: 733–737. DOI: 10.1016 / 0047-2484 (88) -2. [CrossRef] [Google Scholar] 7. Лю В., Мартинон-Торрес М., Цай Ю.Дж., Син С., Тонг Х.В., Пей С.В., Сиер М.Дж., Ву Х.Х., Эдвардс Р.Л., Ченг Х. и др. Самые ранние люди современного типа на юге Китая. Природа. 2015; 526: 696–699. DOI: 10,1038 / природа15696. [PubMed] [CrossRef] [Google Scholar] 8. Хаблин Дж. Дж., Бен-Нсер А., Бейли С. Е., Фрейдлайн С. Е., Нойбауэр С., Скиннер М. М., Бергманн И., Ле Кабек А., Бенацци С., Харвати К. и др.Новые окаменелости из Джебель-Ирхуда, Марокко и панафриканского происхождения Homo sapiens. Природа. 2017; 546: 289–292. DOI: 10,1038 / природа22336. [PubMed] [CrossRef] [Google Scholar] 9. Richter D., Grün R., Joannes-Boyau R., Steele T.E., Amani F., Rué M., Fernandes P., Raynal J.P., Geraads D., Ben-Ncer A., et al. Возраст окаменелостей гомининов из Джебель-Ирхуда, Марокко, и истоки среднего каменного века. Природа. 2017; 546: 293–296. DOI: 10,1038 / природа22335. [PubMed] [CrossRef] [Google Scholar] 10. Гершковиц И., Weber G.W., Quam R., Duval M., Grün R., Kinsley L., Ayalon A., Bar-Matthews M., Valladas H., Mercier N., et al. Самые ранние современные люди за пределами Африки. Наука. 2018; 359: 456–459. DOI: 10.1126 / science.aap8369. [PubMed] [CrossRef] [Google Scholar] 11. Лалуэза-Фокс К., Гилберт М.Т.П. Обзор палеогеномики архаичных гомининов. Curr. Биол. 2011; 21: R1002 – R1009. DOI: 10.1016 / j.cub.2011.11.021. [PubMed] [CrossRef] [Google Scholar] 12. Прюфер К., Расимо Ф., Паттерсон Н., Джей Ф., Санкарараман С., Сойер С., Heinze A., Renaud G., Sudmant P.H., De Filippo C. и др. Полная последовательность генома неандертальца из Горного Алтая. Природа. 2014; 505: 43–49. DOI: 10,1038 / природа12886. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 13. Прюфер К., де Филиппо К., Гроте С., Мафессони Ф., Корлевич П., Хайдиньяк М., Верно Б., Сков Л., Шей П., Пейрегне С. и др. Геном неандертальца с высоким охватом из пещеры Виндия в Хорватии. Наука. 2017; 505: 43–49. DOI: 10.1126 / science.aao1887. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 14.Мондал М., Казальс Ф., Сюй Т., Далл’Олио Г.М., Пибус М., Нетеа М.Г., Комас Д., Лааюни Х., Ли К., Маджумдер П.П. и др. Геномный анализ андаманских островов дает представление о миграции древнего человека в Азию и адаптации. Nat. Genet. 2016; 48: 1066–1070. DOI: 10,1038 / нг. 3621. [PubMed] [CrossRef] [Google Scholar] 15. Се П., Вернер А.Е., Уолл Дж. Д., Лашанс Дж., Тишкофф С.А., Гутенкунст Р.Н., Хаммер М.Ф. Модельный анализ данных полного генома показывает сложную эволюционную историю, включающую архаическую интрогрессию у пигмеев Центральной Африки.Genome Res. 2016; 26: 291–300. DOI: 10.1101 / gr.196634.115. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 16. Медина-Гомес К., Лао О., Риваденейра Ф. Эволюция сложных черт в человеческих популяциях. В: Понтаротти П., редактор. Эволюционная биология: эволюция «я / не-я», эволюция видов и сложных свойств, методы и концепции. Издательство Springer International Publishing; Чам, Швейцария: 2017. С. 165–186. [Google Scholar] 17. Санкарараман С., Паттерсон Н., Ли Х., Пэабо С., Райх Д. Дата скрещивания неандертальцев и современных людей.PLoS Genet. 2012; 8: e1002947. DOI: 10.1371 / journal.pgen.1002947. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 18. Уолл Д.Д., Ян М.А., Джей Ф., Ким С.К., Дюран Э.Ю., Стивисон Л.С., Жинью К., Вернер А., Хаммер М.Ф., Слаткин М. Более высокие уровни неандертальского происхождения у жителей Восточной Азии, чем у европейцев. Генетика. 2013 DOI: 10.1534 / genetics.112.148213. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 19. Фу К., Ли Х., Мурджани П., Джей Ф., Слепченко С. М., Бондарев А.-П., Пяабо С. Последовательность генома 45000-летнего современного человека из Западной Сибири.Природа. 2014; 514: 445–449. DOI: 10,1038 / природа13810. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 20. Сегин-Орландо А., Корнелиуссен Т.С., Сикора М., Маласпинас А.С., Маника А., Мольтке И., Альбрехтсен А., Ко А., Маргарян А., Моисеев В. и др. Геномная структура европейцев насчитывает не менее 36 200 лет. Наука. 2014; 346: 1113–1118. DOI: 10.1126 / science.aaa0114. [PubMed] [CrossRef] [Google Scholar] 21. Санкарараман С., Маллик С., Паттерсон Н., Райх Д. Объединенный ландшафт денисовских и неандертальских предков у современных людей.Curr. Кипятить. 2016; 26: 1241–1247. DOI: 10.1016 / j.cub.2016.03.037. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 22. Грин Р.Э., Краузе Дж., Бриггс А.В., Маричич Т., Стензел У., Кирчер М., Паттерсон Н., Ли Х., Чжай В., Фриц М.Х. и др. Предварительная последовательность генома неандертальца. Наука. 2010; 328: 710–722. DOI: 10.1126 / science.1188021. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 23. Маллик С., Ли Х., Липсон М., Мэтисон И., Гимрек М., Рачимо Ф., Чжао М., Ченнагири Н., Норденфельт С., Тандон А. и др. Проект разнообразия генома Саймонса: 300 геномов из 142 различных популяций. Природа. 2016; 538: 201–206. DOI: 10,1038 / природа18964. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 24. Браунинг С.Р., Чжоу Ю., Туччи С., Эйки Дж.М. Анализ данных о человеческой последовательности выявляет два импульса архаичной денисовской примеси. Клетка. 2018; 178: 53–61. DOI: 10.1016 / j.cell.2018.02.031. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 26. Хаммер М.Ф., Вернер А.Э., Мендес Ф.Л., Уоткинс Дж.К., Уолл Дж. Д. Генетические свидетельства архаической примеси в Африке. Proc. Natl. Акад. Sci. США. 2011; 108: 15123–15128. DOI: 10.1073 / pnas.11017. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 75. Ху Х., Петуси Н., Глусман Г., Ю. Ю., Болендер Р., Таши Т., Дауни Дж. М., Роуч Дж. К., Коул А. М., Лоренцо Ф. Р. и др.Эволюционная история тибетцев, выведенная из полногеномного секвенирования. PLoS Genet. 2017; 13: e1006675. DOI: 10.1371 / journal.pgen.1006675. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 76. Уэрта-Санчес Э., Джин Х., Бьянба З., Питер Б.М., Винкенбош Н., Лян Ю., Йи Х., Хе М., Сомел М., Ни П. и др. Адаптация к высоте у тибетцев, вызванная интрогрессией денисовской ДНК. Природа. 2014; 512: 194–197. DOI: 10,1038 / природа13408. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 77.Таши Т., Скотт Ридинг Н., Вурен Т., Чжан Х., Мур Л.Г., Ху Х., Тан Ф., Шестакова А., Лоренцо Ф., Бурджанивова Т. и др. Повышенная функциональность пролилгидроксилазы EGLN1 (PHD2 D4E: C127S) в сочетании с полиморфизмом EPAS1 (HIF-2α) снижает концентрацию гемоглобина у тибетских горцев. J. Mol. Med. 2017; 95: 665–670. DOI: 10.1007 / s00109-017-1519-3. [PubMed] [CrossRef] [Google Scholar] 78. Куач Х., Кинтана-Мурси Л. Жизнь в адаптивном мире: геномное расчленение рода Homo и его иммунный ответ.J. Exp. Med. 2017; 214: 877–894. DOI: 10.1084 / jem.20161942. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 79. Andrés A.M., Hubisz M.J., Indap A., Torgerson D.G., Degenhardt J.D., Boyko A.R., Gutenkunst R.N., White T.J., Green E.D., Bustamante C.D., et al. Задачи балансирующего отбора в геноме человека. Мол. Биол. Evol. 2009. 26: 2755–2764. DOI: 10,1093 / molbev / msp190. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 80. ДеДжорджио М., Ломюллер К.Е., Нильсен Р. Модельно-ориентированный подход к идентификации сигнатур древнего сбалансированного отбора в генетических данных.PLoS Genet. 2014; 10: e1004561. DOI: 10.1371 / journal.pgen.1004561. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 81. Леффлер Э.М., Гао З., Пфейфер С., Сегурель Л., Аутон А., Венн О., Боуден Р., Бонтроп Р., Уолл Дж. Д., Селла Г. и др. Множественные примеры древнего уравновешивающего отбора, общие для человека и шимпанзе. Наука. 2013; 340: 1578–1582. DOI: 10.1126 / science.1234070. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 82. Норман П.Дж., Аби-Речед Л., Гендзехадзе К., Корбель Д., Gleimer M., Rowley D., Bruno D., Carrington C.V.F., Chandanayingyong D., Chang Y.H. и др. Необычный отбор на рецепторе естественных киллерных клеток KIR3DL1 / S1 у африканцев. Nat. Genet. 2007; 39: 1092–1099. DOI: 10,1038 / нг2111. [PubMed] [CrossRef] [Google Scholar] 83. Сингл Р.М., Мартин М.П., Гао X., Мейер Д., Йегер М., Кидд Дж.Р., Кидд К.К., Кэррингтон М. Глобальное разнообразие и доказательства совместной эволюции KIR и HLA. Nat. Genet. 2007; 39: 1114–1119. DOI: 10,1038 / нг2077. [PubMed] [CrossRef] [Google Scholar] 84.Аби-Рачед Л., Джобин М.Дж., Кулкарни С., Маквинни А., Далва К., Грагерт Л., Бабрзаде Ф., Гаризаде Б., Луо М., Пламмер Ф.А. и др. Формирование иммунной системы современного человека путем смешивания нескольких регионов с архаичными людьми. Наука. 2011; 334: 89–94. DOI: 10.1126 / science.1209202. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 85. Ясукочи Ю., Охаши Дж. Выяснение происхождения аллельной линии HLA-B * 73: выиграли ли современные люди от архаической интрогрессии? Иммуногенетика. 2017; 69: 63–67. DOI: 10.1007 / s00251-016-0952-8. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 87. Пикрелл Дж. Совместный анализ функциональных геномных данных и полногеномных ассоциаций 18 человеческих черт. Являюсь. J. Hum. Genet. 2014; 94: 559–573. DOI: 10.1016 / j.ajhg.2014.03.004. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 88. Шауб М.А., Бойл А.П., Кундаже А., Бацоглу С., Снайдер М. Связывание ассоциаций болезней с регуляторной информацией в геноме человека. Genome Res. 2012; 22: 1748–1759. DOI: 10.1101 / гр.136127.111. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 89. Даннеманн М., Прюфер К., Келсо Дж. Функциональные последствия интрогрессии неандертальцев у современных людей. Genome Biol. 2017; 18:61. DOI: 10.1186 / s13059-017-1181-7. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 90. Sams A.J., Dumaine A., Nédélec Y., Yotova V., Alfieri C., Tanner J.E., Messer P.W., Barreiro L.B. Адаптивно интрогрессированный гаплотип неандертальца в локусе OAS функционально влияет на врожденные иммунные ответы у людей.Genome Biol. 2016; 17: 246. DOI: 10.1186 / s13059-016-1098-6. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 91. Моцци А., Форни Д., Кальяни Р., Поццоли У., Клеричи М., Сирони М. Четкие селективные силы и неандертальская интрогрессия сформировали генетическое разнообразие генов, участвующих в нарушениях развития нервной системы. Sci. Отчет 2017; 7 DOI: 10.1038 / s41598-017-06440-4. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 92. Simonti C.N., Vernot B., Bastarache L., Bottinger E., Carrell D.S., Chisholm R.Л., Кросслин Д. Р., Хеббринг С. Дж., Ярвик Г. П., Кулло И. Дж. И др. Фенотипическое наследие смешения современных людей и неандертальцев. Наука. 2016; 351: 737–741. DOI: 10.1126 / science.aad2149. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 93. Таскент Р.О., Алиоглу Н.Д., Фер Э., Мелике Донертас Х., Сомел М., Гоккумен О. Вариации и функциональное влияние неандертальцев в Западной Азии. Геномный фурункул. Evol. 2017; 9: 3516–3524. DOI: 10.1093 / GBE / evx216. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 94.Баррейро Л.Б., Кинтана-Мурси Л. От эволюционной генетики до иммунологии человека: как отбор формирует гены защиты хозяина. Nat. Преподобный Жене. 2010; 11: 17–30. DOI: 10,1038 / NRG2698. [PubMed] [CrossRef] [Google Scholar] 95. Бринкворт Дж. Ф., Баррейро Л. Б. Вклад естественного отбора в сегодняшнюю восприимчивость к хроническим воспалительным и аутоиммунным заболеваниям. Curr. Opin. Иммунол. 2014; 31: 66–78. DOI: 10.1016 / j.coi.2014.09.008. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 96. Корбетт С., Куртиоль А., Луммаа В., Мурад Дж., Стернс С. Переход к современности и хронические заболевания: несоответствие и естественный отбор. Nat. Преподобный Жене. 2018: 1–12. DOI: 10.1038 / s41576-018-0012-3. [PubMed] [CrossRef] [Google Scholar] 97. Сирони М., Клеричи М. Гипотеза гигиены: эволюционная перспектива. Микробы заражают. 2010; 12: 421–427. DOI: 10.1016 / j.micinf.2010.02.002. [PubMed] [CrossRef] [Google Scholar] 99. Хант К.А., Жернакова А., Тернер Г., Хип Г.А.Р., Франке Л., Брюненберг М., Романос Дж., Динесен Л.К., Райан А.В., Панесар Д. и др. Недавно идентифицированные варианты генетического риска целиакии, связанные с иммунным ответом. Nat. Genet. 2008. 40: 395–402. DOI: 10,1038 / нг.102. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 100. Жернакова А., Эльберс К.С., Ферверда Б., Романос Дж., Трынка Г., Дюбуа П.С., де Ковель К.Г.Ф., Франке Л., Остинг М., Барисани Д. и др. Эволюционный и функциональный анализ локусов глютенового риска выявил Sh3B3 как защитный фактор против бактериальной инфекции. Являюсь. Дж.Гм. Genet. 2010; 86: 970–977. DOI: 10.1016 / j.ajhg.2010.05.004. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 101. Даннеманн М., Андрес А.М., Келсо Дж. Интрогрессия неандертальских и денисовских гаплотипов способствует адаптивной изменчивости Toll-подобных рецепторов человека. Являюсь. J. Hum. Genet. 2016; 98: 22–33. DOI: 10.1016 / j.ajhg.2015.11.015. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 103. Агует Ф., Браун А.А., Кастель С.Э., Дэвис Дж.Р., Хе Й., Джо Б., Мохаммади П., Парк Ю.С., Парсана П., Сегре А.В. и др. Генетические эффекты на экспрессию генов в тканях человека. Природа. 2017; 550: 204–213. DOI: 10,1038 / природа24277. [CrossRef] [Google Scholar] 104. Ардли К.Г., ДеЛука Д.С., Сегре А.В., Салливан Т.Дж., Янг Т.Р., Гельфанд Э.Т., Троубридж К.А., Маллер Дж.Б., Тукиайнен Т., Лек М. и др. Пилотный анализ экспрессии генотипа и ткани (GTEx): регуляция многотканевых генов у людей. Наука. 2015; 348: 648–660. DOI: 10.1126 / science.1262110. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 105.Филд Й., Бойл Е.А., Телис Н., Гао З., Голтон К.Дж., Голан Д., Йенго Л., Рошело Г., Фрогель П., Маккарти М.И. и др. Обнаружение адаптации человека за последние 2000 лет. Наука. 2016; 354: 760–764. DOI: 10.1126 / science.aag0776. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 106. Тишкофф С.А., Рид Ф.А., Ранчиаро А., Войт Б.Ф., Бэббит С.С., Сильверман Дж. С., Пауэлл К., Мортенсен Х.М., Хирбо Дж. Б., Осман М. и др. Конвергентная адаптация персистенции лактазы человека в Африке и Европе.Nat. Genet. 2007 г. DOI: 10.1038 / ng1946. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Перевод в англо-немецком словаре LEO
Aktivieren Sie JavaScript für mehr Features und höhere Geschwindigkeit beim Abfragen.
Прилагательные :: Существительные :: Аналогичные :: Связанные :: Обсуждения :: 9000 9000 adj5Прилагательные / Наречия | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| архаичный прил. | altertümlich | |||||||||
| 9000 adj5 | 9000 9000 9000 adj5 | archaisch | ||||||||
| | 3 | 3 | 2392 adj5 + | + | + | veraltet + | ||||
существительных | |||||||
|---|---|---|---|---|---|---|---|
| + | + | Архаический период [ИСТОЧНИК] | die Archaik [Археология] 93599359259 | ||||
| 35 | 3 | 359 похожие слова | |||||
| Archaik | |||||||
Обсуждения на форуме, содержащие поисковый запрос | ||
|---|---|---|
| ….. архаичная форма ….. | 2 ответа | |
| Kräuterfrau (ARCHAIC) | Последнее сообщение 22 ноя 12, 12:54 | |
| Hallo, ich suche den altenglischen Begriff für Kräuterfrauen. Und zwar meine ich damit die F… | 9 Ответов | |
| архаический инстинкт | Последнее сообщение 17 января, 19:58 | |
| 75 | ||
| 99 | ||
| Последнее сообщение 10 янв 07, 16:42 | ||
| englischsprachige Quellen siehe zum Beispiel unter http: // www.thefreelibrary.com/bs.asp?ss=t… | 6 ответов | |
| shoon [архаичный] — Schuhe | Последнее сообщение 10 апр 05, 00:32 | |
| http: / /www.bartleby.com/61/82/S0358200.html shoon СУЩЕСТВИТЕЛЬНОЕ: Архаичный Множественное число «обувь». The… | 2 Ответа | |
| , поэтому [архаичный] — warum | Последнее сообщение 11 мая 06, 21:50 | |
| Несокращенный словарь Harper Collins: wherefores warum, weswegen Cassell ‘… | 2 ответа | |
| gloam [архаичный] — Zwielicht | Последнее сообщение 04 декабря 14, 11:57 | |
| http: // www.merriam-webster.com/dictionary/gloam gloam имя существительное Определение GLOAM архаичный:… | 6 ответов | |
| Urbild — архаичное изображение | Последнее сообщение 17 января 05, 16:20 | |
| 2 ответа | ||
| antiquated — altertümelnd | Последнее сообщение 17 января 12, 14:02 | |
| % http://www.dict.cc/?s=altert C3% BCmelnd http: // dict.tu-chemnitz.de/deutsch-englisch/altert%… | 8 Ответов | |
| bizarre —archaic — skurril | Последнее сообщение 02 мая 08, 19:43 | |
| 3 ответа | ||
Другие действия
Узнать больше
Нужна языковая консультация? Обратитесь за помощью к другим пользователей на наших форумах.Редактируйте списки слов
Sortieren Sie Ihre gespeicherten Vokabeln.История поиска
Sehen Sie sich Ihre letzten Suchanfragen an.Английский ⇔ немецкий словарь — leo.org: Начальная страница
SUCHWORT — Перевод в англо-немецком словаре LEO
LEO.org: Ваш онлайн-словарь для англо-немецких переводов. Предлагаем форумы, словарный инструктор и языковые курсы. Также доступно как приложение!
Выучите перевод слова SUCHWORT в англо-немецком словаре LEO.С таблицами существительных / глаголов для различных падежей и времен ✓ ссылки на звуковое произношение и соответствующие обсуждения на форуме ✓ бесплатный тренажер словарного запаса ✓
Запись сохранена в трейнере. Добавить в список слов сейчас?
Ваш вклад был отправлен на форум.
Пространственные корни политики и общества »(онлайн-конференция) — Классическая ассоциация
Мы рады пригласить вас на Международную конференцию «Аттика от позднего бронзового века до конца архаического периода: пространственные корни политики и общества», организованную Университетом Париж 1 Пантеон-Сорбонна и итальянским Археологическая школа в Афинах.
Мероприятие состоится через Zoom в четверг и пятницу, 10 и 11 июня 2021 года, . Программа здесь ниже.
Обратите внимание, что все временные интервалы указаны по афинскому времени (GMT +3).
Конференция открытая и бесплатная, но для посещения мероприятия необходимо предварительное бронирование. Убедительно просим всех желающих зарегистрироваться здесь , или связавшись с организаторами, Николаосом Арванитисом ([электронная почта защищена]) или Аленом Дюплуи ([электронная почта защищена]).Ссылки Zoom и аннотации статей будут разосланы зарегистрированным участникам до начала мероприятия.
Аттика от позднего бронзового века до конца архаического периода
Пространственные корни политики и общества
Международная онлайн-конференция
10–11 июня 2021 года
Парижский университет 1 Пантеон-Сорбонна и итальянская археологическая школа в Афинах
Организаторы: Николаос Арванитис и Ален Дюплуи
В 1964 году Пьер Видаль-Наке и Пьер Левек представили новаторский подход к истории Афин в своей книге Clisthène l’Athénien .Несмотря на низкое почтение древних афинян к Клисфену, два французских историка сделали его «великим архитектором» классических Афин, геометром новой концепции гражданского пространства и времени. Подзаголовок их книги, переведенный на английский только тридцать лет спустя в рамках празднования 2500-летия зарождения демократии, ясно выражает тесную связь, существовавшую между обществом, политикой и космосом в Древней Греции: An Essay on the Изображение пространства и времени в греческой политической мысли с конца шестого века до смерти Платона (1996).С тех пор образ Клисфена изменился, и его собственная роль в политических преобразованиях позднеархического афинского общества была уменьшена, в то время как были разработаны другие интерпретации клисфенических реформ, в том числе идея самоорганизующегося сообщества на часть афинского демоса, которая могла быть частью более широкой региональной тенденции с отголосками в современной Эретрии и Коринфе (Ismard 2010). Тем не менее космос остается важным инструментом интерпретации в древней истории.Этот регион c. 2 550 км2 — это одна из самых хорошо задокументированных территорий древнегреческого города. Соответственно, эта конференция рассматривает использование космических и сетевых теорий в качестве потенциально эвристических инструментов в текущих археологических и исторических исследованиях Древней Греции. Из-за обилия археологических и текстовых свидетельств Аттика представляет собой привилегированную эвристическую обсерваторию социальных идентичностей, выраженных в различных пространственных контекстах от позднего бронзового века до классического периода.Однако многие аспекты доклассических исторических процессов остаются плохо изученными из-за отсутствия прочной связи с критически оцененными пространственными параметрами. Среди географов, историков, социальных теоретиков, антропологов и философов растет осознание конституирующего значения места и пространства, места и ситуации, местности и территориальности, которые изучаются в рамках многомасштабного подхода и как узлы сети взаимодействий. Пространственный подход предполагается даже для тех локусов, реальных или символических, которые можно восстановить в текстовых нарративах: они обладают важной символической и репрезентативной значимостью, в конечном итоге заякоренной в палимпсесте многослойных дискурсов, которые, как таковые, занимают «место» как ментальные / повествовательные проекции.Несмотря на нынешний расцвет ИТ-технологий в археологической практике, большая часть современных ученых по-прежнему не в своей тарелке, когда дело доходит до контекстной визуализации социальных моделей. С другой стороны, при изучении только на основе текстовых свидетельств политическая и социальная динамика не часто объединяет многомерные траектории, которые могут выявить археологические исследования. Мы предполагаем, что социальная динамика и историческое развитие могут быть изучены только тогда, когда динамика самого пространства понимается в четырехмерной перспективе как серия взаимосвязанных событий, передающих действительность.Пространство следует исследовать как физическое выражение когнитивных схем, которые являются культурно специфическими, контекстуальными и историческими. Другими словами, цель конференции — сосредоточить внимание на пространственности исторических процессов и сетей в древней Аттике и на взаимосвязанных масштабах — временном и пространственном, — в которых эти сети перемещаются. В конечном итоге конференция направлена на то, чтобы подвести черту под значимой биографией Аттики на протяжении длительного периода перемен.
Все временные интервалы указаны по афинскому времени (GMT + 3)
Четверг, 10 июня 2021 г.
10:00 — 10:15 — Прием
10:15 — 10:30 — Открытие, Эмануэле Папи, приветственная речь
10:30 — 10:50 — Николаос Арванитис и Ален Дюплуи (Парижский университет 1 Пантеон-Сорбонна), Введение.
Сессия 1 — Аттика от позднего бронзового века до геометрического периода (председатель Элени Андрику)
10:50 — 11:20 — Николас Пападимитриу (Гейдельбергский университет), микенская Аттика: сколько «микенского» и сколько «аттического»?
11:20 — 11:50 — Костас Калогеропулос (Афинская академия), разделенный рекой Эразинос. Северная и Южная равнины Мезогеи от позднего бронзового века до раннего железного века.
11:50 — 12:00 — Перерыв на кофе
12:00 — 12:30 — Флорис ван ден Эйнде (Утрехтский университет), Между Полисом и Этносом.Сетевой подход к развитию Афинского Полиса.
12:30 — 13:00 — Максимилиан Рённберг (Немецкий археологический институт), «Множество вариантов: изменение предпочтений и местных особенностей в размещении захоронений в Аттике от позднего элладского периода до архаического».
13:00 — 13:30 — Николаос Арванитис (Университет Париж 1 Пантеон-Сорбонна), Большие объекты в Аттике за пределами Афин. Дело Элевсина и Ахарная.
13:30 — 14:00 — Обсуждение
14:00 — 16:00 — Перерыв на обед
Сессия 2 — Аттика от геометрического до классического периода (кафедра Анастасия Гадолоу)
16:00 — 16:30 — Николаос Арванитис, Ален Дюплуи, Анастасия Струсопулу (Парижский университет 1 Пантеон-Сорбонна), Анагирус.Анатомия чердачного района.
16:30 — 17:00 — Джеймс Уитли (Университет Кардиффа), От кубков до кратеров: поверхности письма в ранней Аттике (800–500 гг. До н. Э.).
17:00 — 17:10 — Перерыв на кофе
17:10 — 17:40 — Эрих Кистлер (Университет Инсбрука), Чердак Джин между Евномией и Стазисом. Создание альтернативы Athenaion Politeia.
17:40 — 18:10 — Риккардо Ди Чезаре (Университет Фоджи и Итальянская археологическая школа в Афинах), Архаическая Аттика и символическое строительство космоса.Достопримечательности, легенды и культы.
18:10 — 18:40 — Обсуждение
Пятница, 11 июня 2021 г.
Сессия 3 — Афины и их район перед Клисфеном (председатель Кристина Меркури)
10:30 — 11:00 — Тонио Хёльшер (Гейдельбергский университет), архаическая агора Афин. Его расположение в топографической системе Афин.
11:00 — 11:30 — Аннарита Доронцио (Мартин-Лютер-Университет Галле-Виттенберг), ατά κώμας.Исследуемая динамика афинского поселения.
11:30 — 11:40 — Перерыв на кофе
11:40 — 12:10 — Винченцо Капоццоли (Парижский университет 1 Пантеон-Сорбонна), Так называемые городские демы Афин в архаическую эпоху. К изложению свидетельств.
12:10 — 12:40 — Александра Александриду (Университет Янины), Регионализм в Афинах раннего железного века: Внутреннее ядро в Академии.
12:40 — 13:10 — Обсуждение
13:10 — 15:30 — Перерыв на обед
Сессия 4 — Методологические, хронологические и региональные сравнения (председатель Стелла Хрисулаки)
15:30 — 16:00 — Андреас Капетаниос (Ионический университет), Из космоса в Топос.Археология преемственности и изменений в структуре ландшафта Лавриона.
16:00 — 16:30 — Эмери Фаринетти (Итальянская археологическая школа в Афинах), За пределами Аттики: ландшафтные подходы в Беотии и Мегариде.
16:30 — 17:00 — Паоло Карафа (Римский университет «Ла Сапиенца»), Политика, люди и пейзажи. От Аттики до древнего Лацио.
17:00 — 17:20 — Обсуждение
17:20 — 17:30 — Кофе-брейк
Сессия 5 — Перспективы
17:30 — 18:00 — Современные и новые инструменты в Афинах и Аттике, с Николаосом Арванитисом, Аннаритой Доронцио, Аленом Дюплуи, Барборой Вайссовой.
18:00 — 19:30 — Круглый стол с Тонио Хёльшером (Гейдельбергский университет), Ирен Лемос (Оксфордский университет), Робином Осборном (Кембриджский университет), Франсуа де Полиньяком (EPHE, Париж).
Анализ полногеномной последовательности панафриканского набора образцов показывает архаичный поток генов из вымершей основной популяции современных людей в популяции к югу от Сахары | Genome Biology
Ingman M, Kaessmann H, Paabo S, Gyllensten U.Вариации митохондриального генома и происхождение современного человека. Природа. 2000; 408: 708–13.
CAS Статья PubMed Google Scholar
Гарриган Д., Хаммер М.Ф. Реконструкция происхождения человека в эпоху генома. Nat Rev Genet. 2006; 7: 669–80.
CAS Статья PubMed Google Scholar
Кэмпбелл М.С., Хирбо Дж. Б., Таунсенд Дж. П., Тишкофф С.А. Заселение африканского континента и диаспоры в новый мир.Curr Opin Genet Dev. 2014; 29: 120–32.
CAS Статья PubMed PubMed Central Google Scholar
Hublin JJ, Ben-Ncer A, Bailey SE, Freidline SE, Neubauer S, Skinner MM, Bergmann I, Le Cabec A, Benazzi S, Harvati K, Gunz P. Новые окаменелости из Джебель-Ирхуда, Марокко и панафриканское происхождение Homo sapiens. Природа. 2017; 546: 289–92.
CAS Статья PubMed Google Scholar
Филипсон DW. Африканская археология. 3-е изд. Кембридж: Издательство Кембриджского университета; 2005.
Книга. Google Scholar
Хенн Б.М., Жинью С.Р., Джобен М., Гранка Дж. М., Макферсон Дж. М., Кидд Дж. М., Родригес-Ботиг Л., Рамачандран С., Хон Л., Брисбин А. и др. Геномное разнообразие охотников-собирателей предполагает южноафриканское происхождение современного человека. Proc Natl Acad Sci U S. A. 2011; 108: 5154–62.
CAS Статья PubMed PubMed Central Google Scholar
Пикрелл Дж. К., Паттерсон Н., Барбьери С., Бертольд Ф., Герлах Л., Гульдеманн Т., Куре Б., Мполока С. В., Накагава Х., Науманн С. и др. Генетическая предыстория юга Африки. Nat Commun. 2012; 3: 1143.
Артикул PubMed PubMed Central Google Scholar
Schlebusch CM, Skoglund P, Sjodin P, Gattepaille LM, Hernandez D, Jay F, Li S, De Jongh M, Singleton A, Blum MG, et al. Геномные вариации в семи группах кхоэ-сан показывают адаптацию и сложную африканскую историю.Наука. 2012; 338: 374–9.
CAS Статья PubMed Google Scholar
Richter D, Grun R, Joannes-Boyau R, Steele TE, Amani F, Rue M, Fernandes P, Raynal JP, Geraads D, Ben-Ncer A, et al. Возраст окаменелостей гомининов из Джебель-Ирхуда, Марокко, и истоки среднего каменного века. Природа. 2017; 546: 293–6.
CAS Статья PubMed Google Scholar
Diamond J, Bellwood P. Фермеры и их языки: первые дополнения. Наука. 2003; 300: 597–603.
CAS Статья PubMed Google Scholar
Бехар Д.М., Виллемс Р., Судьял Х., Блю-Смит Дж., Перейра Л., Мецпалу Е., Скоццари Р., Маккан Х., Цур С., Комас Д. и др. Рассвет человеческого матрилинейного разнообразия. Am J Hum Genet. 2008; 82: 1130–40.
CAS Статья PubMed PubMed Central Google Scholar
Li JZ, Absher DM, Tang H, Southwick AM, Casto AM, Ramachandran S, Cann HM, Barsh GS, Feldman M, Cavalli-Sforza LL, Myers RM. Человеческие взаимоотношения во всем мире, выведенные из полногеномных моделей изменчивости. Наука. 2008; 319: 1100–4.
CAS Статья PubMed PubMed Central Google Scholar
Тишкофф С.А., Рид Ф.А., Фридлендер Ф.Р., Эрет К., Ранчиаро А., Фромент А., Хирбо Дж. Б., Авомойи А. А., Бодо Дж. М., Думбо О. и др.Генетическая структура и история африканцев и афроамериканцев. Наука. 2009; 324: 1035–44.
CAS Статья PubMed PubMed Central Google Scholar
Gronau I, Hubisz MJ, Gulko B, Danko CG, Siepel A. Байесовский вывод демографии древнего человека на основе индивидуальных последовательностей генома. Нат Жене. 2011; 43: 1031–4.
CAS Статья PubMed PubMed Central Google Scholar
Veeramah KR, Wegmann D, Woerner A, Mendez FL, Watkins JC, Destro-Bisol G, Soodyall H, Louie L, Hammer MF. Раннее расхождение предков KhoeSan от предков других современных людей подтверждается основанным на ABC анализом данных аутосомного ресеквенирования. Mol Biol Evol. 2012; 29: 617–30.
CAS Статья PubMed Google Scholar
Найт А., Андерхилл П.А., Мортенсен Х.М., Животовский Л.А., Лин А.А., Хенн Б.М., Луис Д., Рулен М., Маунтин Дж. Л..Дивергенция африканской Y-хромосомы и мтДНК дает представление об истории языков щелчков. Curr Biol. 2003; 13: 464–73.
CAS Статья PubMed Google Scholar
Животовский Л.А., Розенберг Н.А., Фельдман М.В. Особенности эволюции и распространения современного человека, полученные на основе полногеномных микросателлитных маркеров. Am J Hum Genet. 2003. 72: 1171–86.
CAS Статья PubMed PubMed Central Google Scholar
Kim HL, Ratan A, Perry GH, Montenegro A, Miller W, Schuster SC. Койсанские охотники-собиратели были самой большой популяцией на протяжении большей части истории современного человечества. Nat Commun. 2014; 5: 5692.
CAS Статья PubMed PubMed Central Google Scholar
Верду П., Дестро-Бисол Г. Африканские пигмеи, что стоит за именем? Hum Biol. 2012; 84: 1–10.
Артикул PubMed PubMed Central Google Scholar
Verdu P, Becker NS, Froment A, Georges M, Grugni V, Quintana-Murci L, Hombert JM, Van der Veen L, Le Bomin S, Bahuchet S, et al. Социокультурное поведение, смешение по признаку пола и эффективный размер популяции пигмеев и непигмеев Центральной Африки. Mol Biol Evol. 2013; 30: 918–37.
CAS Статья PubMed PubMed Central Google Scholar
Археология, язык и африканское прошлое. Роджер Бленч. Лэнхэм: AltaMira Press; 2006 г.п. 388. ISBN 978-0-7591-0466-2 (мягкая обложка).
Патин Э, Лаваль Дж., Баррейро Л. Б., Салас А., Семино О, Сантачиара-Бенеречетти С., Кидд К. К., Кидд Дж. Р., Ван дер Вин Л., Хомберт Дж. М. и др. Выявление демографической истории африканских фермеров и охотников-собирателей карликов с использованием набора данных многократного повторного упорядочивания. PLoS Genet. 2009; 5: e1000448.
Артикул PubMed PubMed Central Google Scholar
Верду П., Аустерлиц Ф., Эступ А., Виталис Р., Жорж М., Тери С., Фромент А., Ле Бомин С., Гессен А., Хомберт Дж. М. и др.Происхождение и генетическое разнообразие пигмеев-охотников-собирателей из Западной Центральной Африки. Curr Biol. 2009; 19: 312–8.
CAS Статья PubMed Google Scholar
Batini C, Lopes J, Behar DM, Calafell F, Jorde LB, van der Veen L, Quintana-Murci L, Spedini G, Destro-Bisol G, Comas D. Анализ демографической истории африканских пигмеев из полных митохондриальных геномов. Mol Biol Evol. 2011; 28: 1099–110.
CAS Статья PubMed Google Scholar
Шей П., Вирама К.Р., Лашанс Дж., Тишкофф С.А., Уолл Д.Д., Хаммер М.Ф., Гутенкунст Р.Н. Анализ полногеномной последовательности охотников-собирателей пигмеев из Западной Центральной Африки раскрывает сложную демографическую историю и идентифицирует гены-кандидаты в условиях положительного естественного отбора. Genome Res. 2016; 26: 279–90.
CAS Статья PubMed PubMed Central Google Scholar
Патин Э, Лопес М., Гроллемунд Р., Верду П., Хармант С., Квач Х, Лаваль Г., Перри Г. Х., Баррейро Л. Б., Фромент А. и др.Расселение и генетическая адаптация говорящих на банту популяций в Африке и Северной Америке. Наука. 2017; 356: 543–6.
CAS Статья PubMed Google Scholar
Петерсен Д.К., Либигер О., Тиндалл Е.А., Харди Р.А., Ханник Л.И., Глэшофф Р.Х., Мукерджи М., Индийский консорциум вариаций генома, Фернандес П., Хааке В. и др.: Сложные образцы геномной примеси в южной Африке. PLoS Genet 2013, 9: e1003309.
CAS Статья PubMed PubMed Central Google Scholar
Гурдасани Д., Карстенсен Т., Текола-Айеле Ф., Пагани Л., Тачмазиду И., Хатзикотулас К., Картикеян С., Ильес Л., Поллард М. О., Чоудхури А. и др. Проект изменения африканского генома формирует медицинскую генетику в Африке. Природа. 2015; 517: 327–32.
CAS Статья PubMed Google Scholar
Пикрелл Дж., Паттерсон Н., Ло П.Р., Липсон М., Бергер Б., Стоункинг М., Пакендорф Б., Рейх Д. Древние западно-евразийские предки в южной и восточной Африке.Proc Natl Acad Sci U S. A. 2014; 111: 2632–7.
CAS Статья PubMed PubMed Central Google Scholar
Грин RE, Краузе Дж., Бриггс А.В., Маричич Т., Стензель У., Кирхер М., Паттерсон Н., Ли Х., Чжай В., Фриц М.Х. и др. Предварительная последовательность генома неандертальца. Наука. 2010; 328: 710–22.
CAS Статья PubMed PubMed Central Google Scholar
Райх Д., Грин Р. Э., Кирчер М., Краузе Дж., Паттерсон Н., Дюран Э. Ю., Виола Б., Бриггс А. В., Стензель Ю., Джонсон П. Л. и др. Генетическая история архаической группы гомининов из Денисовой пещеры в Сибири. Природа. 2010; 468: 1053–60.
CAS Статья PubMed PubMed Central Google Scholar
Sanchez-Quinto F, Botigue LR, Civit S, Arenas C, Avila-Arcos MC, Bustamante CD, Comas D, Lalueza-Fox C. Популяции Северной Африки несут в себе след неандертальцев.PLoS One. 2012; 7: e47765.
CAS Статья PubMed PubMed Central Google Scholar
Henn BM, Botigue LR, Gravel S, Wang W, Brisbin A, Byrnes JK, Fadhlaoui-Zid K, Zalloua PA, Moreno-Estrada A, Bertranpetit J, et al. Геномное происхождение североафриканцев поддерживает обратную миграцию в Африку. PLoS Genet. 2012; 8: e1002397.
CAS Статья PubMed PubMed Central Google Scholar
Арауна Л. Р., Мендоса-Ревилья Дж., Мас-Сандовал А., Изаабель Х, Бекада А., Бенхамамуш С., Фадхлауи-Зид К., Заллуа П., Хелленталь Г., Комас Д. Недавние исторические миграции сформировали генофонд арабов и берберов в Северная Африка. Mol Biol Evol. 2017; 34: 318–29.
CAS PubMed Google Scholar
Ван С., Лашанс Дж., Тишкофф С.А., Эй Дж., Син Дж. Очевидные вариации в составе неандертальцев среди африканских популяций согласуются с потоком генов из неафриканских популяций.Genome Biol Evol. 2013; 5: 2075–81.
Артикул PubMed PubMed Central Google Scholar
Пруфер К., Расимо Ф., Паттерсон Н., Джей Ф., Санкарараман С., Сойер С., Хайнце А., Рено Г., Судмант PH, де Филиппо С. и др. Полная последовательность генома неандертальца из Горного Алтая. Природа. 2014; 505: 43–9.
Артикул PubMed Google Scholar
Plagnol V, Wall JD. Возможная предковая структура в человеческих популяциях. PLoS Genet. 2006; 2: e105.
Артикул PubMed PubMed Central Google Scholar
Hammer MF, Woerner AE, Mendez FL, Watkins JC, Wall JD. Генетические свидетельства архаической примеси в Африке. Proc Natl Acad Sci U S. A. 2011; 108: 15123–8.
CAS Статья PubMed PubMed Central Google Scholar
Лашанс Дж., Верно Б., Эльберс С.К., Ферверда Б., Фромент А., Бодо Дж. М., Лема G, Фу В., Ньямбо Т. Б., Реббек Т. Р. и др. История эволюции и адаптация к последовательностям полного генома различных африканских охотников-собирателей. Клетка. 2012; 150: 457–69.
CAS Статья PubMed PubMed Central Google Scholar
Xu D, Pavlidis P, Taskent RO, Alachiotis N, Flanagan C, DeGiorgio M, Blekhman R, Ruhl S, Gokcumen O.Архаическая интрогрессия гомининов в Африке вносит вклад в функциональную генетическую изменчивость MUC7 слюны. Mol Biol Evol. 2017; 34: 2704–15.
CAS Статья PubMed PubMed Central Google Scholar
Хсие П., Вернер А.Е., Уолл Д.Д., Лашанс Дж., Тишкофф С.А., Гутенкунст Р.Н., Хаммер М.Ф. Модельный анализ данных полного генома показывает сложную эволюционную историю, включающую архаическую интрогрессию у пигмеев Центральной Африки.Genome Res. 2016; 26: 291–300.
CAS Статья PubMed PubMed Central Google Scholar
Дурвасула А., Санкарараман С. Восстановление сигналов призрачной архаической примеси в геномах современных африканцев. bioRxiv. 2018. https://doi.org/10.1101/285734.
Скоглунд П., Томпсон Дж. С., Прендергаст М. Е., Миттник А., Сирак К., Хайдинджак М., Сали Т., Роланд Н., Маллик С., Пельцер А. и др. Реконструкция доисторической структуры населения Африки.Клетка. 2017; 171: 59–71 e21.
CAS Статья PubMed PubMed Central Google Scholar
Kuhlwilm M, Gronau I, Hubisz MJ, de Filippo C, Prado-Martinez J, Kircher M, Fu Q, Burbano HA, Lalueza-Fox C, de la Rasilla M, et al. Древние гены перетекают от ранних современных людей к восточным неандертальцам. Природа. 2016; 530: 429–33.
CAS Статья PubMed PubMed Central Google Scholar
Posth C, Wissing C, Kitagawa K, Pagani L, van Holstein L, Racimo F, Wehrberger K, Conard NJ, Kind CJ, Bocherens H, Krause J. Глубоко дивергентный архаический митохондриальный геном обеспечивает нижнюю временную границу для потока африканских генов в неандертальцев . Nat Commun. 2017; 8: 16046.
CAS Статья PubMed PubMed Central Google Scholar
Таваре С., Болдинг Д. Д., Гриффитс Р. К., Доннелли П. Вывод времени слияния на основании данных последовательности ДНК.Генетика. 1997. 145: 505–18.
CAS PubMed PubMed Central Google Scholar
Берторелле Дж., Бенаццо А., Мона С. ABC как гибкая система оценки демографии в пространстве и времени: некоторые минусы, много плюсов. Mol Ecol. 2010; 19: 2609–25.
CAS Статья PubMed Google Scholar
Sisson SA, Fan Y, Beaumont M. Справочник по приближенным байесовским вычислениям: CRC Press, Taylor & Francis Group; 2018.ISBN 1439881502, 9781439881507.
Jiang B, Wu T-Y, Zheng C, Wong WH. Сводная статистика обучения для приблизительного байесовского вычисления с помощью глубокой нейронной сети. Стат Sin. 2017; 27: 1595–618.
Google Scholar
Mondal M, Bertranpetit J, Lao O. Приближенные байесовские вычисления с глубоким обучением поддерживают третью архаичную интрогрессию в Азии и Океании. Nat Commun. 2019; 10: 246.
Артикул PubMed PubMed Central Google Scholar
Gutenkunst RN, Hernandez RD, Williamson SH, Bustamante CD. Вывод совместной демографической истории нескольких популяций из многомерных данных частоты SNP. PLoS Genet. 2009; 5: e1000695.
Артикул PubMed PubMed Central Google Scholar
Schuster SC, Miller W., Ratan A, Tomsho LP, Giardine B, Kasson LR, Harris RS, Petersen DC, Zhao F, Qi J, et al. Полные геномы койсанов и банту из южной Африки.Природа. 2010; 463: 943–7.
CAS Статья PubMed PubMed Central Google Scholar
Чоудхури А., Рамзи М., Хейзелхерст С., Арон С., Бардиен С., Бота Г., Чимуса Е. Р., Кристоффельс А., Гамильдиен Дж., Сефид-Дашти М. Дж. И др. Секвенирование всего генома для лучшего понимания генетической изменчивости среди южноафриканцев. Nat Commun. 2017; 8: 2062.
Артикул PubMed PubMed Central Google Scholar
Пембертон Т.Дж., Абшер Д., Фельдман М.В., Майерс Р.М., Розенберг Н.А., Ли Дж.З. Геномные паттерны гомозиготности в человеческих популяциях во всем мире. Am J Hum Genet. 2012; 91: 275–92.
CAS Статья PubMed PubMed Central Google Scholar
Rosenberg MS. Коррелограмма пеленга: новый метод анализа направленной пространственной автокорреляции. Геогр Анал. 2000. 32: 267–78.
Артикул Google Scholar
Джей Ф, Сйодин П, Якобссон М, Блюм МГ. Анизотропная изоляция расстоянием: основные направления генетической дифференциации человека. Mol Biol Evol. 2013; 30: 513–25.
CAS Статья PubMed Google Scholar
Александр Д.Х., Новембре Дж., Ланге К. Быстрая модельная оценка происхождения неродственных особей. Genome Res. 2009; 19: 1655–64.
CAS Статья PubMed PubMed Central Google Scholar
Афган Э, Бейкер Д., ван ден Бик М., Бланкенберг Д., Бувье Д., Чех М., Чилтон Дж., Клементс Д., Кораор Н., Эберхард С. и др. Платформа Galaxy для доступных, воспроизводимых и совместных биомедицинских анализов: обновление 2016 г. Nucleic Acids Res. 2016; 44: W3 – W10.
CAS Статья PubMed PubMed Central Google Scholar
Паттерсон Н., Мурджани П., Луо И., Маллик С., Роланд Н., Жан И., Геншорек Т., Вебстер Т., Райх Д.Древняя примесь в истории человечества. Генетика. 2012; 192: 1065–93.
Артикул PubMed PubMed Central Google Scholar
Ли Х., Дурбин Р. Вывод истории человеческой популяции на основе отдельных полногеномных последовательностей. Природа. 2011; 475: 493–6.
CAS Статья PubMed PubMed Central Google Scholar
Нахман М.В., Кроуэлл С.Л. Оценка скорости мутации на нуклеотид у человека.Генетика. 2000; 156: 297–304.
CAS PubMed PubMed Central Google Scholar
Конрад Д.Ф., Киблер Д.Е., ДеПристо М.А., Линдси С.Дж., Чжан Ю., Казальс Ф., Идагдур Ю., Хартл К.Л., Торроха С., Гаримелла К.В. и др. Различия в частоте мутаций по всему геному внутри и между человеческими семьями. Нат Жене. 2011; 43: 712–4.
CAS Статья PubMed PubMed Central Google Scholar
Кэмпбелл С.Д., Чонг Дж. Х., Малиг М., Ко А., Дюмон Б. Л., Хан Л., Вивес Л., О’Роак Б. Дж., Судмант П. Х., Шендуре Дж. И др. Оценка частоты мутаций человека с использованием аутозиготности в популяции-основателе. Нат Жене. 2012; 44: 1277–81.
CAS Статья PubMed PubMed Central Google Scholar
Марнер Л., Ньенгаард Дж. Р., Тан Й., Паккенберг Б. Заметная потеря миелинизированных нервных волокон в мозге человека с возрастом. J Comp Neurol. 2003. 462: 144–52.
Артикул PubMed Google Scholar
Даннеманн М., Расимо Ф. Что-то старое, что-то заимствованное: примесь и адаптация в эволюции человека. Curr Opin Genet Dev. 2018; 53: 1–8.
CAS Статья PubMed Google Scholar
Гилкс В., Ричардсон С., Шпигельхальтер Д. Марковская цепь Монте-Карло на практике. Тейлор и Фрэнсис; 1995. ISBN: 9780412055515.
Excoffier L, Estoup A, Cornuet JM. Байесовский анализ модели примеси с мутациями и произвольно связанными маркерами. Генетика. 2005; 169: 1727–38.
CAS Статья PubMed PubMed Central Google Scholar
Beltrame MH, Rubel MA, Tishkoff SA. Выводы африканской эволюционной истории из геномных данных. Curr Opin Genet Dev. 2016; 41: 159–66.
CAS Статья PubMed PubMed Central Google Scholar
Campana MG, Бауэр Массачусетс, Крэбтри П.Дж. Древняя ДНК для археолога: будущее африканских исследований. Afr Archaeol Rev.2013; 30: 21–37.
Артикул Google Scholar
Li H, Durbin R. Быстрое и точное согласование коротких считываний с помощью преобразования Барроуза-Уиллера. Биоинформатика. 2009; 25: 1754–60.
CAS Статья PubMed PubMed Central Google Scholar
ДеПристо М.А., Бэнкс Э., Поплин Р., Гаримелла К.В., Магуайр Дж. Р., Хартл С., Филиппакис А.А., дель Анхель Г., Ривас М.А., Ханна М. и др. Структура для обнаружения вариаций и генотипирования с использованием данных секвенирования ДНК следующего поколения. Нат Жене. 2011; 43: 491–8.
CAS Статья PubMed PubMed Central Google Scholar
Карбон Л., Харрис Р.А., Гнерре С., Вирама К.Р., Лоренте-Галдос Б., Хаддлстон Дж., Мейер Т.Дж., Эрреро Дж., Роос К., Акен Б. и др.Геном гиббона и быстрая эволюция кариотипа мелких обезьян. Природа. 2014; 513: 195–201.
CAS Статья PubMed PubMed Central Google Scholar
Бехар Д.М., Ван Овен М., Россет С., Мецпалу М., Лугвали Е.Л., Сильва Н.М., Кивисилд Т., Торрони А., Виллемс Р. «Коперниканская» переоценка митохондриального дерева ДНК человека от его корня. Am J Hum Genet. 2012; 90: 675–84.
CAS Статья PubMed PubMed Central Google Scholar
Донмез Н., Брудно М. Хапсемблер: ассемблер для высокополиморфных геномов. В: Bafna V, Sahinalp SC, ред. Исследования в области вычислительной молекулярной биологии; 2011 //; Берлин, Гейдельберг. Гейдельберг: Springer Berlin; 2011. с. 38–52.
Глава Google Scholar
Wei W, Ayub Q, Chen Y, McCarthy S, Hou Y, Carbone I, Xue Y, Tyler-Smith C. Калиброванная филогения Y-хромосомы человека, основанная на повторном секвенировании. Genome Res. 2013; 23: 388–95.
CAS Статья PubMed PubMed Central Google Scholar
Скалецкий Х., Курода-Кавагути Т., Минкс П.Дж., Кордум Х.С., Хиллиер Л., Браун Л.Г., Реппинг С., Пынтикова Т., Али Дж., Биери Т. и др. Специфическая для мужчин область Y-хромосомы человека представляет собой мозаику классов дискретных последовательностей. Природа. 2003; 423: 825–37.
CAS Статья PubMed PubMed Central Google Scholar
Van Geystelen A, Decorte R, Larmuseau MH. AMY-tree: алгоритм использования полногеномного SNP, вызывающий филогенетические приложения Y-хромосомы. BMC Genomics. 2013; 14: 101.
Артикул PubMed PubMed Central Google Scholar
Ван Ч., Шпих З.А., Дегнан Дж. Х., Якобссон М., Пембертон Т. Дж., Харди Дж. А., Синглтон А. Б., Розенберг Н. А.. Сравнение пространственных карт популяционно-генетической изменчивости человека с использованием анализа Прокруста.Stat Appl Genet Mol Biol. 2010; 9: Статья 13.
PubMed Google Scholar
Розенберг М.С., Андерсон CD. ПАССАЖ: Анализ закономерностей, пространственная статистика и географическая экзегеза. Версия 2. Методы Ecol Evol. 2011; 2: 229–32.
Артикул Google Scholar
Kunsch HR. Складной нож и бутстрап для общих стационарных наблюдений. Ann Stat. 1989; 17: 1217–41.
Артикул Google Scholar
Excoffier L, Foll M. fastsimcoal: непрерывный симулятор геномного разнообразия при произвольно сложных эволюционных сценариях. Биоинформатика. 2011; 27: 1332–4.
CAS Статья PubMed Google Scholar
Мейер М., Кирхер М., Гансоге М.Т., Ли Х., Расимо Ф., Маллик С., Шрайбер Дж. Г., Джей Ф., Пруфер К., де Филиппо С. и др.Последовательность генома с высоким охватом архаичного денисовца. Наука. 2012; 338: 222–6.
CAS Статья PubMed PubMed Central Google Scholar
Геродот и источники истории архаического периода
Этой главой «Темный век» оставлен позади, и начинается так называемый архаический период греческой истории (примерно с 800 г. до н.э. до персидского Войны). В начале этого периода греки начали основывать колонии по всему Средиземноморью, и это движение привело к значительному расширению ареала греческой цивилизации.К счастью, это тоже в период
.История Греции: 1300–30 гг. До н.э., первое издание. Виктор Паркер.
© Виктор Паркер, 2014 г. Опубликовано в 2014 году компанией John Wiley & Sons, Ltd.
Это период, когда снова начинается письменная традиция, которая позволяет обсуждать эти далеко идущие события более подробно, чем в случае «Темного века».
Краткое введение в главный литературный источник архаического периода, Истории Геродота, будет поэтому особенно полезным для установления того, что Геродот, хотя и писал во второй половине пятого века до нашей эры, все же мог перечислить много ценных сведений о восьмом веке до нашей эры. и седьмой век.Рассказы Геродота в высшей степени читабельны и занимательны, хотя его работа в целом преследует серьезную цель. Вот два программных утверждения из работы:
Ниже приводится «история» Геродота Галикарнасского, что человеческие дела не могут исчезнуть с течением времени; эти великие и чудесные дела, которые признают как греки, так и не греки, могут быть более широко известны. . . (из произведения) [Я буду рассматривать] как малые, так и большие человеческие города, ибо из тех, которые были великими давным-давно, многие теперь малы; но некоторые из них, которые в мои дни были великими, раньше были маленькими.А потому, зная, что среди людей процветание никогда не пребывает долго на одном и том же месте, я буду говорить об обоих в равной степени. (HDT I 5)
Хотя основная часть работ Геродота посвящена конфликтам между греками и персами в течение первых двух десятилетий пятого века, он все еще может многое сказать о людях и событиях шестого века, но постепенно меньше о седьмом и восьмом веке. Это подтверждает общую достоверность информации; ибо этого следует ожидать от любого, кто честно собирает информацию — больше информации о более ранних периодах, чем о более поздних, вызовет скептицизм (см. Ephorus, BNJ 70, Fr.9).
Тем не менее, ученые, которые в последние годы обсуждали использование Геродотом устных источников, часто приходили к удручающим выводам относительно достоверности информации, которую он предлагает для раннего периода. Это обсуждение, хотя и чрезвычайно полезное, иногда основывается на неправильном восприятии, а именно, что Геродот работал только с устными источниками, а не с устно изложенными традициями, отредактированными и упорядоченными в соответствии с другими письменными источниками, которые были доступны еще с середины седьмого века.
Геродот, по сути, провел тщательный обзор ранее существовавшей греческой литературы на предмет исторической информации, которая могла бы помочь ему при сортировке устных преданий. Например, он упоминает Гигеса, исторического царя Лидии в середине седьмого века до нашей эры (о Лидийском царстве см. Кратко главу 9). Этот Гигес независимо засвидетельствован в надписях Ашшурбанипала, царя Ассирии (668 — около 627 г. до н.э.). Геродот не мог знать о последнем, но у него тоже было письменное свидетельство Гигеса в стихотворении Архилоха (фр.19 West) — см. Hdt. I 12. Из этого Геродот сделал вывод, что Архилох и Гигес были современниками. В качестве дополнительного свидетельства общей надежности Геродота он заявляет (I 15), что киммерийцы вторглись в Лидию примерно в это время. Надписи ассирийского царя Ашшурбанипала — несмотря на различные отличия от рассказа Геродота — подтверждают тот же факт.1
Более важный пример того, как Геродот использовал письменный источник в качестве руководства, встречается в V 95, когда он цитирует Алкея Митиленского (фр.428 Лобель-Пейдж) как источник войны с участием Писистрата Афинского, Питтака Митиленского и Периандра Коринфского (эти тираны шестого века см. В главе 7). Неясно, какая часть рассказа происходит непосредственно от Алкея, но даже если из стихотворения Алкея следует отсылка к тому, что произошло с Алкеем в битве между афинянами и митиленцами, Геродот все же прочитал поэзию Алкея и признал, что Алкей описал события которые были такими же, как и в ряде устных преданий об этой войне.
Геродот также читал стихи Сафо, современницы Алкея (см. Алкей, фр. 384 Лобель-Пейдж), которая высмеивала своего брата Харакса за его отношения с легендарной куртизанкой Родопидой во времена Амасиса, исторического царя Египта ( 570-526 гг. До н.э .; HDT. II 135 = Сафо, фр. 202 Лобель-Пейдж). Читая стихи Солона (см. Главу 8), Геродот заметил, что Солон в начале шестого века посетил кипрский город Саламин, когда его более поздний царь Филокипр был молодым человеком (Hdt.V 113 = Солон, о. 19 Запад). Этот Филокипр был отцом участника Ионического восстания (499–494 гг. До н.э .; см. Главу 9). Во всем этом можно увидеть, как Геродот использует письменные источники в качестве исторических свидетельств, чтобы понять многие устные традиции, которые он собрал.
, благодаря его усердию, а также усилиям других авторов, таких как Антиох Сиракузский (конец пятого века — см. Также вставку 5.2), колонизаторское движение в основном происходит в полном свете истории.
Прославление архаики | Автор: Сет Бенардете,
С публикацией в 1903 г. сборника досократиков Диля, идея независимого величия «архаического» периода греческой литературы (ок.650-450 до н.э.), отличные как от эпического периода Гомера и Гесиода, так и от классической эпохи, вступили в свои права. Архаическая эпоха Сафо, Пиндара и Архилоха была не только признана отдельной традицией, но и считалась олицетворением особых и достойных восхищения ценностей в искусстве, литературе и философии. Это было также на рубеже веков, когда архаическая скульптура была впервые оценена, и репутация классических скульпторов, таких как Фидий (пятый век) и скульпторов-натуралистов, таких как Пракситель (четвертый век), пострадала соразмерно.
Герман Френкель, профессор классики в Стэнфорде с 1935 года, отчасти разделяет эту точку зрения на архаическую эпоху и придает ей новое значение. Когда его книга «Ранняя греческая поэзия и философия » была впервые опубликована на немецком языке Американской филологической ассоциацией в 1951 году, она сразу же вызвала восхищение у классиков. (Недавний перевод на английский язык был последним проектом Моисея Хадаса перед его смертью.) Книгу Френкеля считали важной не только потому, что он смог составить непрерывное повествование из фрагментарных записей архаического периода, но и потому, что он утверждал, что обнаружил в ней что-то особенное. последовательный образ мыслей.
Архаический период, по его мнению, отличался несентиментальным реализмом, в отличие от «романтизма» эпического периода. В то время как в «Илиаде » человек является «полностью частью своего мира», архаические поэты, как считает Френкель, открыли «внутреннюю суть личной жизни». Но Фрэнкель намеревается сделать больше, чем доказать разделение эпической и архаической мысли. Он восхищается архаичными ценностями. «Литературные памятники архаической эпохи, — пишет он, — открыли нам способ существования, который по-своему завершен и способен придать смысл жизни и смерти человека.Это один из самых благородных первобытных образов и образов человечества, которые нам известны ».
Суть книги Френкеля — анализ лирической поэзии от Архилоха до Пиндара и философии от Фалеса до Гераклита. Ни в коем случае не пренебрегая ими, он обсуждает Гесиода и Гомера в основном потому, что они представляют идеи, отличные от идей архаических поэтов и философов, то есть как свидетельство предлагаемой им прерывности. Древние говорили о Гомере как о «поэте». Френкель потребовал бы этот титул для Пиндара, написавшего «автономные стихи», столь же ярких, как сама жизнь, и более удовлетворяющих.Для Френкеля Пиндара — завершение архаического искусства и доказательство разрыва между архаическим и классическим менталитетом, поскольку едва ли какие-либо ценности, которые отстаивал Пиндар, пережили его. После него, пишет Френкель, «немела целая эпоха».
Френкель отнюдь не написал общепринятую историю раннегреческой литературы. Эсхил, современник Пиндара, не упоминается, а Вакхилид привлекает к себе лишь поверхностное внимание из-за того, что Френкель в высшей степени озабочен своим тезисом: архаический период Греции следует понимать как таковой, а не как предшественник классической эпохи.Существует определенная несогласованность между мастерским анализом свидетельств Френкелем и его более грубым тезисом о том, что составляло единство архаического периода. Для Френкеля тексты — «документы своего возраста», а художники олицетворяют эту эпоху. Таким образом, Пиндар «был и оставался архаичным греком», а Эсхил и другие его современники — нет.
Попытка Френкеля различить единство архаической эпохи опирается на две взаимодополняющие идеи: идеи полярности и самости. Под полярностью Френкель имеет в виду идею, которую, по его мнению, внесла архаическая мысль, о том, что «качества могут быть созданы только в том случае, если [их] противоположности возникли в одно и то же время.«Древние греки выражали мысли и чувства в противоположностях. Как следствие, пишет Френкель, архаический период видел «противоположные силы, неизбежные в одном и том же» и «жизнь как последовательность чередований между противоположными крайностями», в то время как в философии «космическая структура и космический процесс [интерпретировались] как взаимодействие противоположностей ». 1
Рассказ Френкеля об архаическом «я», хотя и гораздо более подробный, напоминает книгу Бруно Снелла « Discovery of Mind » (1947).Снелл предположил, что именно в архаичную эпоху было открыто «я» как отдельная реальность. Под этим Френкель и Снелл подразумевают, что именно в архаической лирической поэзии человек обнаружил, что у него есть душа, и что его чувства не были полностью вопросом его физических функций. Эти чувства берут начало в человеке, но ни в коем случае не принадлежат исключительно ему. Человек изолирован, но он также типичен. «Греческие лирические поэты, — пишет Френкель, — не стремились заинтересовать себя своей особой чувствительностью, а скорее стремились продемонстрировать общее и основное на собственном примере.Как Френкель пытается объединить эти две идеи полярности и себя, лучше всего можно увидеть в его обсуждении поэта Архилоха.
Архилох, согласно Френкелю, основал архаическую эпоху. Его поэзия — это здесь, сейчас и индивидуальное эго; это как его первичные данные, так и окончательные данные, «без оговорок и без сомнения». Чтобы продемонстрировать свои утверждения об автономности архаической эпохи, Френкель складывает два фрагмента из Архилоха: «лиса знает многое, а еж — одно большое» (фр.103) и «… одно важное дело, которое я умею делать — ужасно расплачиваться печалью за причиненное мне горе» (фр. 66). Согласно Френкелю, Архилох — это еж, и «впервые в европейской литературе эго становится полярной противоположностью не-эго. Самость, существованию которой угрожает распад и разрушение из-за признания «эфемерной» природы человека, утверждает свое собственное существо путем конфликта и защиты от других ».
Но разве Архилох — ёжик? В другом фрагменте (фр.1) Архилох говорит, что знает две вещи: «Я оба, последователь бога Эниалиоса [Ареса], и я понимаю искусство, которое является даром Муз». Фрэнкель не считает, что одна из вещей, которые может знать лис, — это то, как выглядеть как ёжик. Более того, Архилох не допускает иронии. Фрэнкель цитирует фрагмент 22:
Меня совершенно не волнуют золотые богатства Гигеса: я не испытываю к нему зависти. То, как ведут себя боги, меня не трогает; трон тирана я не хочу, потому что ничего этого не вижу перед моими глазами …Но когда я вижу, как такой-то и такой-то проходит мимо меня в гордости и силе своих грязно нажитых денег, тогда все, что я хочу, — это бросить топор ему в голову. Так сказал Харон, плотник с Тасоса.
Френкель не сомневается, что Архилох — это Харон. Для Френкеля нравственное возмущение Харона не могло быть скрытой формой зависти. Архилох просто хотел показать, что «блеск высочайшего» мало для него значит по сравнению со скромным успехом человека, которого он ненавидит. «Мировая история меркнет перед тем, что происходит на нашей собственной улице.
Далее, хотя большая часть его поэзии состоит из инвектив, из этого не обязательно следует — как предполагает Френкель, — что Архилох считал себя оппозиционным Гомеру. Поскольку гнев неотделим от чувства несправедливости, мы не знаем, просто ли Архилох исследовал одну из тем Илиады года , самое первое слово которой — «гнев».
Согласно Френкелю, поэзия Архилоха в чистом виде очерчивает «мгновенное состояние индивидуального эго».Но это кажется неубедительным, поскольку Архилох действительно представлял себя в других обличьях (например, в облике плотника Харона). Для Френкеля индивидуальное эго является в то же время репрезентативным или типичным эго — он отмечает, что «ранняя лирика целиком или почти полностью связана с разговором с кем-то». Но это подрывает его взгляд на архаическую полярность; поскольку, если «я» всегда коррелирует с «вами», они образуют «мы», и оппозиция между эго и не-я исчезает. Если Архилоха можно рассматривать как репрезентативное эго, то в тезисе Френкеля о природе архаического менталитета есть неразрешенное противоречие.
В своей попытке отделить архаическую эпоху как от того, что ей предшествовало, так и от того, что последовало за ней, Френкель иногда вынужден прибегать к другим несоответствиям, которые — даже если они несущественны — сбивают с толку. Он обращается к единству архаической эпохи — которая, в конце концов, является его собственным построением — как к стандарту для интерпретации сохранившихся текстов и фрагментов. Одним из результатов построения Френкеля является то, что каждый поэт или философ перестает быть интересным сам по себе. Его возраст всегда «глубже», чем он есть.Следовательно, философ Анаксимандр — «хороший архаический мыслитель», а «добро», очевидно, модифицирует архаику, а не мыслителя; то есть Анаксимандр мыслит архаичной личностью.
Взывая к духу эпохи, Френкель также стремится минимизировать трудность интерпретации того, что, в конце концов, в значительной степени является фрагментами. В начале периода, описываемого Френкелем, у нас есть четыре полных (хотя и не без истолкования) стихотворения — Гомер Илиада и Одиссея и Гесиода Теогония и Работы и дни .В конце — «Оды Победы» Пиндара. Между ними у нас есть одно полное стихотворение Сафо, несколько второстепенных произведений и фрагменты. Мы не знаем, писали ли какие-нибудь поэты книги в период между Гесиодом и аттическими трагедиями. (Было бы очень важно, если бы мы знали, потому что в книге паузы между стихами являются частью речи поэта и должны учитываться, когда мы пытаемся их интерпретировать.) Френкель, хотя хорошо осведомлен о состоянии доказательств. , настаивает на использовании этих произведений и фрагментов в качестве иллюстраций более крупной сущности, архаической лирики.
Трудность, которая возникает из-за того, что Фрэнкель использует доказательства, становится очевидной при чтении им дошедшего до нас целого стихотворения Сафо. Поэма Сафо адресована Афродите. Она молится, чтобы Афродита пришла ей на помощь, как и раньше; Затем она вспоминает, как Афродита однажды спросила Сафо, кто обидел ее, и пообещала, что скоро все изменится, и возлюбленный станет любовником. Сафо, наконец, повторяет свою молитву и просит Афродиту прийти в качестве ее военного союзника.Поэма показывает, как богиня любви может быть нелюбимой; страдающий любовник переживает безответную любовь как несправедливость и поэтому смотрит на собственные последующие страдания возлюбленного из-за любви как на наказание, справедливость которого гарантирует бог. Эта мораль проникает в, казалось бы, самые аморальные переживания, и это наше отражение в стихотворении. Но мы никогда не узнаем, что Сапфо хотел, чтобы мы из этого сделали.
Фрэнкель, однако, уверенно читает стихотворение: «Сафо пережила то, что мы должны назвать внутренним событием, переход от мучительных мук к уверенной надежде, которая позже осуществилась, что она поняла и передала словами, соответствующими ее вере в божественная сила любви.«Чтение Френкеля, конечно, может быть правильным. Но нельзя не задаться вопросом, не интересует ли его Сапфо больше как иллюстрация определенного аспекта архаической лирики, чем ее интерпретация в свете самой себя.
В одном редком случае диссертация Френкеля заставляет его неправильно переводить фрагмент Сапфо. Фрагмент начинается словами «Он кажется мне равным богам», а ближе к концу говорит: «Кажется, я почти мертв» (фр. 31). Френкель передает это так: «Он кажется равным богам… почти я мертв.Опустив «мне» в начале и «мне кажется» в конце, Френкель может сказать о стихотворении, что «все стоит на одном и том же уровне. Никакие глубины души не открываются ». Чтобы Сафо оставалась простой и прямой, он упускает искусно подразумеваемый контраст между иллюзией бессмертия Сафо в другом человеке и ее иллюзией собственной смерти.
Превосходство Френкеля заключается больше в его обсуждении стилей мышления и письма, чем в его анализе мысли и написанного.(Это еще более очевидно в его сборнике эссе « Wege und Formen frühgriechischen Denkens », который составляет техническую основу «Ранняя греческая поэзия и философия ».) Поразительно, что он посвящает не более одного-двух предложений этому. сюжеты Илиады и Одиссеи , но исследуют гомеровского человека и его богов подробно. То, что он говорит о них, почти всегда поучительно, и есть много наблюдений, которых больше нигде в литературе не найти.Но Френкель несколько преувеличивает различие между поэтом и его материалом: для него материал не полностью находится под контролем поэта и может быть понят сам по себе как документ эпохи.
Френкель пишет, например, что «зрелое и позднее эпическое искусство Илиады , обязанное своим совершенством гуманизации и духовному очищению, прославляет более дикие и грубые типы человечества, которые оно превзошло». Он собирает все это из одного отрывка, в котором Нестор, пытаясь унять гнев Ахилла и Агамемнона, вспоминает, как его слушали даже самые жестокие герои прошлого.Но обращение Нестора к разуму, по словам Френкеля, не может быть «подлинно эпическим», поскольку Илиада прославляет жестокость ушедшей эпохи. «Нестор с его советами по дружескому примирению олицетворяет современную разумность». Таким образом, для Френкеля подлинной является только жестокость стихотворения; его человечность предвещает развращение эпической формы. Он не может допустить, чтобы Илиада была морально двусмысленной в своих намерениях.
Проведенное Френкелем различие между подлинной жестокостью и развращающим разумом в Илиаде кажется искусственным; 2 когда он это делает, мы можем видеть, что он поддерживает особое чтение греческой литературы, которое само имеет свою историю.В то время как он описывает Архилоха как поэта нового реализма, Френкель также предполагает, что в «Архилохе» «Человечество шагает вперед, как оно есть, в героической наготе; с мужественной решимостью он лишает себя всех ограничивающих условностей и всяких надуманных украшений ». Похоже, что сам Френкель, если не Архилох, был виновен в том самом «романтическом прославлении существования», которое он находит у Гомера. Подобно тому, как энтузиазм Френкеля по поводу архаического периода напоминает тенденции конца девятнадцатого века в немецкой классической науке, он до сих пор читает Илиаду года так, как ее читали в то время, между франко-прусской войной и Первой мировой войной. , когда гомеровская наука была на пике, и jeunesse dorée Европы нашли в Илиаде славное оправдание войны, которую они все более и более тосковали.И снова в книге Френкеля есть что-то старомодное по духу, даже несмотря на то, что она актуальна. Он игнорирует бесстрастный взгляд Илиады на войну — на честь, которую она дает, а также на зверство, которое она навязывает.
Не исключено, что Френкель видит в «Теогонии » Гесиода начало философии. «В этих стихах воплощены глубокие онтологические рассуждения», — утверждает он, утверждая, что Теогония «включает размышления по основным вопросам метафизики.Форма Theogony отчасти является «мифическим покровом», под которым выражается гораздо больше. Френкель называет Theogony неорганизованным стихотворением, но оно не является неорганизованным, и он не сформулировал адекватно метафизику, которую он в ней обнаруживает.
Даже краткое изложение теогонии (буквально порождение или рождение богов) может предложить, почему и как эта работа имеет структуру, поскольку она описывает борьбу между Ураном (небом) — первым повелителем вселенной — его дети титанов и их дети олимпийцы.
В начале генеалогий лежат не только Хаос и Земля, но и прекрасный Эрос, который в одинаковой мере овладевает умами людей и богов. Земля производит Уран и соединяется с ним; но Уран ненавидит своих детей-титанов и безуспешно пытается изгнать их в недра Земли. Они свергают его, и их лидер, Кронос, кастрирует своего отца, когда Уран лежит на вершине Земли. Затем Кронос и Титаны правят вселенной. Таким образом, разделение неба и земли происходит из-за ненависти Урана к своим детям.Небеса становятся средними и, следовательно, постоянными. Когда Афродита рождается — из морской пены, куда Кронос бросил гениталии своего отца, — Эрос заражается местью.
Олимпийцы, дети титанов, восстают и — под предводительством сына Кроноса Зевса — изгоняют их в подземный мир, где правит брат Зевса Аид. Царствование Зевса представляет собой окончательное устройство вселенной. Там, где Уран не смог изгнать титанов, Зевсу это удалось. Он восстанавливает красоту любви, создавая всемогущую иллюзию Пандоры, и (хотя Пандора также является наказанием за кражу огня Прометеем) примиряет человечество с поколением своей красотой.Затем Зевс заменяет якобы постоянное разделение неба и земли добровольным разделением — Атлас разделяет их и может прекратить делать это в любой момент. Таким образом, люди также примиряются с поколением через страх. И ужас, и красота делают их довольными своей участью.
Теогония описывает порядок, который Зевс налагает на людей, а также его организацию божественного царства, где, хотя и не самый старый из богов, он заставляет более старых божеств вписаться в свой новый порядок.В генеалогической истории богов Гесиод видит телеологию, конечную цель, раскрытую в организации вселенной Зевсом. Более того, в интерпретации происхождения вселенной в Теогонии есть теодицея — подтверждение божественного порядка, служащего оправданием пути богов к человечеству. Гесиод показывает, как с правлением Зевса разум и любовь наконец работают вместе. Браки Зевса (его первый — до Метида , ремесло, мать Афины) символизируют порядок, который он приносит.
Но если Гесиода следует рассматривать как протометафизика, то сначала необходимо разобраться в странной истории Теогонии , а не, как это делает Френкель, выбирать те фразы и отрывки, которые предполагают спекулятивное понимание бытия и небытие. Френкель утверждает, что обсуждение Гесиода бытия и небытия выражает архаичную концепцию полярных противоположностей. «Все в бытии, — пишет он об отрывке из Теогонии , — существует благодаря тому факту, что ему противостоит (пространственно, временно и логически) пустое небытие.Гесиод, согласно Френкелю, противопоставляет положительную систему земли, моря и неба отрицательной системе пустоты, Тартара и ночи. Но то, что сделал здесь Френкель, — это стереть различие между поэтической парой Ночь и День и философской парой Один и Многие, то есть различие между эстетическим и ноэтическим феноменом, под рубрикой полярных противоположностей. Таким образом, Френкелю становится слишком легко утверждать, что поэты и философы «согласны».
Ибо, когда он обсуждает истоки философии в архаическую эпоху, Френкель оказывается в ловушке своей темы.И Гесиод, и Гераклит олицетворяют «поляризованный образ мышления архаической эпохи». Следовательно, «то, что начал Гесиод, Гераклит довел до исполнения». Френкеля не беспокоит тот факт, что Гесиод был поэтом, а Гераклит философом, поскольку он считает, что до появления философии в шестом веке «философские мысли могли быть определены только в форме мифа». Хотя Френкель действительно признает разницу между поэзией и философией, его концепция единства архаической эпохи не позволяет ему увидеть несводимость различия.Френкель рассматривает миф как одно «средство» архаической мысли, лирическую поэзию как другое, а философию как третье, причем все три представляют для него дух архаической эпохи.
Но есть разница между поэтами и философами. Философы предполагают, что человеческий разум сам по себе компетентен, в то время как поэты полагаются на муз, которые однажды сказали Гесиоду: «Мы знаем, как лгать, как правда, и мы знаем, когда захотим, чтобы раскрыть правду». Музы, насколько нам известно, исключительно греки; никто другой не признал, что их единственный доступ к знанию богов был через поэзию.В Гесиоде единственные боги, которые индивидуально называются олимпийскими, — это Зевс и музы. Поскольку музы являются таким же препятствием, как и путь к познанию, мудрость поэтов столь же неоднозначна, как и их боги. Кажется, что он стоит где-то между общепринятой и философской мудростью и, следовательно, в конечном итоге зависит от того, что ниже или выше, чем он сам.
Любитель мифов, говорит Аристотель, только в некотором смысле любит мудрость. Френкель не принял во внимание критику поэзии Аристотелем и, как следствие, упустил истинный смысл философской революции.Если Гомер заставил Геру сказать, что Океан является источником всего сущего, а Фалес говорит, что вода является принципом всего сущего, то Фалес не просто «демифологизировал» Гомера, как сказал бы Френкель. Для Гомера Океан — начало, но не конец всего; генеалогии богов открыты и никогда не возвращаются к себе, чтобы окружить мир. Напротив, Фалес, по-видимому, был первым, кто предположил, что сумма вещей образует понятное целое; Из чего, через что и за что были одним и тем же.
Сглаживание Френкелем различия между поэзией и философией приводит его к аналогичной ошибке при обсуждении Гесиода. В то время как для Гесиода существование отдельных и различимых поколений или генеалогий является фундаментальным, все ранние философы отрицают порождение, считая, что ничто не возникает или не имеет отдельной природы. Досократиков интересовало не становление, а бытие. Точно так же Земля для поэтов является наивысшим или одним из высших богов; ни один из философов никогда не позволял земле быть самой по себе принципом всех вещей.
Философия отличается от мифологии, потому что она довольствуется объяснением не только незнакомого и пугающего, но также знакомого и утешительного. По сути, это шага Fränkel, антиантропоморфный. Жизнь не является поводом для удивления у Гесиода; смерть есть. В мифе солнечное затмение обычно означает, что этого пожелал бог; но для досократической философии причина солнечного затмения основана на причине его сияния. Здесь Френкель столкнулся с главной трудностью в интерпретации греческой литературы и философии: с природой богов.Римский энциклопедист Варрон проводил различие между богами города, богами поэтов и богами философов. Поскольку Френкель не исходит из этого полезного различия, он никогда не сможет выйти за рамки общей «религиозности» поэтов и философов. Он во всем руководствуется двумя вещами: пониманием морали, которое находит «хорошее» и «красивое» двусмысленным, но не «справедливым», и волей к очарованию, которая наиболее явно проявляется в его восхвалении Пиндара.
Греки изобрели поэзию и открыли философию.Гомер учил поэтов лгать, и Фалес первым предложил узнать правду обо всем. Философы спорят, поэты представляют. Однако поэты были раньше философов. Сознательное подражание тому, что есть, кажется, на века предшествовало сомнению в том, что есть. Кроме того, поэты скрывались до того, как философы осознали, что должно было быть открыто, и все философы отрицали, что красота и ложь породили истину. Несмотря на красноречивые утверждения Френкеля об обратном, философы, похоже, ничем не обязаны поэтам.
Историческая проблема происхождения философии, однако, менее важна, чем существенный вопрос как для поэзии, так и для философии: статус мудрости. Поэты первыми назвали себя мудрыми, и философам так и не удалось отобрать у них этот титул. Ссора между поэзией и философией, которая теперь известна в своей наиболее вырожденной форме как проблема двух культур, впервые была сформулирована в период между Гомером и Пиндаром. Это самая глубокая тема Раннегреческой поэзии и философии .