Фотосинтез. Темновая фаза
Автор статьи Чергинцев Д.А.
В предыдущей статье мы рассмотрели физиологические процессы, происходящие в листе в ходе так называемой световой фазы фотосинтеза, в которой при непосредственном участии квантов солнечного света происходит образование восстановленных соединений (NADPH+H+) и ATP. И ATP и NADPH+H+ будут в дальнейшем использованы в различных процессах и метаболических путях, в основном – при фиксации атмосферного СО2, а так же – в ассимиляции нитрата и аммония из почвы. На рисунке 1 схематично представлена связь между световой и темновой фазами фотосинтеза. Закаченные в тилакоид в точках сопряжения (цитохром b6/f комплекс и марганцевый кластер) протоны подкисляют люмен и вместе с этим происходит подщелачивание стромы хлоропласта. Далее по градиенту концентрации (в сторону меньшей концентрации) протоны выходят через канал, образованный Fo субъединицей АТФ-синтазы и создаваемое протонами механическое вращения Fo передается на статор (F1), где происходит катализ образования ATP. Электроны, выбитые из P700 PSI, восстанавливают ферредоксин, который, в свою очередь, в окислительно-восстановительной реакции, катализируемой ферредоксин-НАДФ
В темновой фазе происходят процессы, напрямую с солнечным светом не связанные, но большинство ключевых ферментов, работающих на этой стадии, активны только на свету. Зависимая от света работа ферментов определяется несколькими регуляторными механизмами: pH стромы, наличие двухвалентных катионов (преимущественно Mg2+), а также и в основном благодаря взаимодействию с тиоредоксином. Тиоредоксины – группа белков, имеющаяся у всех живых организмов, представители которой отвечают за сигнализацию в ответ на изменение окислительно-восстановительного (redox) потенциала клетки. Тиоредоксины имеют довольно специфическую укладку и два расположенных рядом остатка цистеина. В окисленной форме цистеины образуют дисульфидный мостик (Cys-S-S-Cys), в восстановленной – две тиольные (сульфгидрильные) группы (2Cys-SH). Окисляясь или восстанавливаясь сами, тиоредоксины могут, соответственно, восстанавливать или окислять другие субстраты. Восстанавливая дисульфидные связи в других белках, цистеины регулируют их активность и, помимо этого, могут поставлять электроны для различных redox реакций. Восстановление тиоредоксинов может проходить разными способами. В частности, тиоредоксин может восстанавливаться электронами с восстановленного ферредоксина при участии ферредоксин-тиоредоксинредуктазы (см рис. 2). Таким образом, происходит передача прямого сигнала от электрон-транспортной цепи (ЭТЦ) тилакоида к ферментам различных реакций и осуществляется непосредственный контроль последних. Очевидно, что данная система функционирует только при работе ЭТЦ, то есть – на свету. Помимо этого тиоредоксины могут восстанавливаться с помощью тиоредоксинредуктаз, принимая электроны от NADPH+H
На рисунке 3 схематично изображены основные участники темновой фазы фотосинтеза, подверженные регуляции с помощью тиоредоксиновой системы, хотя, несомненно, регуляция гораздо более обширная. На свету восстановленный благодаря работе ЭТЦ хлоропласта пул тиоредоксинов активирует работу ферментов, отвечающих за путь фиксации CO2 и синтеза глюкозы, в то время как активность ферментов путей анаэробного дыхания – окисления глюкозы (гликолиз и пентозофосфатный путь), будут ингибирована. В темноте ситуация противоположная: тиоредоксиновый пул весь окислен, восстановить его нечем, ферменты пути фиксации углерода не активируются, зато протекают пути окисления синтезированной ранее на свету глюкозы. Помимо тиоредоксиновой регуляции, как уже было сказано, наблюдается регуляция с помощью pH. Те же ферменты, что активируются днем тиоредоксинами, увеличивают свою активность и благодаря повышенным дневным значениям pH стромы хлоропласта (pH=8.0).
Рис 3. Светозависимая регуляция компонентов темновой фазы фотосинтеза.
Непосредственно фиксация углекислого газа, то есть – превращение углерода из окисленной неорганической формы в восстановленную органическую, происходит в восстановительном пентозофосфатном цикле (пути) или иначе — цикле Кальвина. Допустимое сокращение – ВПФП. Путь, который правильнее называть с учетом всех исследователей циклом Кальвина — Бенсона — Бассама (Calvin–Benson–Basshamcycle, CBB), был открыт с использованием СО2 с изотопом 14C. Стадии цикла были выяснены с помощью анализа последовательности включения 14C в разные углеводные продукты. В 1961 году за открытие цикла трем исследователям была присуждена Нобелевская премия по химии.
Цикл Кальвина состоит из трех стадий: 1) карбоксилирование; 2) восстановление; 3) регенерация.
Стадия карбоксилирования состоит из одной реакции, катализируемой рибулозобисфосфат карбоксилазой-оксигеназой или RuBisCO. Пожалуй, RuBisCO – самый распространенный фермент на Земле и при этом очень древний. Содержание RuBisCO может составлять порядка 50% массы растворимых белков зеленых листьев, при этом концентрация фермента на порядки выше концентрации субстрата – СО2, что абсолютно нехарактерно для ферментативных реакций. У растений, большинства водорослей и фотосинтетических бактерий RuBisCO состоит из 8 больших субъединиц (L, 55 kDa), кодируемых пластидным геномом и 8 малых субъединиц (S, 13 kDa), кодируемых ядерным геномом. Большие субъединицы являются каталитическими, малые – регуляторными, и они не являются необходимыми для самой реакции карбоксилирования. L субъединицы организованы в димеры, при этом активные сайты карбоксилирования образованы аминокислотами обеих частей димера, таким образом, RuBisCO с формулой L8S8 содержит 8 активных сайтов (бывают и другие типы RuBisCO).
Схема строения активного центра RuBisCO представлена на рисунке 4. Для осуществления реакции карбоксилирования рибулозо-1,5-бисфосфата (RuBP) требуется первоначальная активация и самого активного центра (рисунок 5). RuBisCO проявляет активность только в том случае, когда ε-аминогруппа лизина в 201 положении ковалентно связана с СО2. После присоединения СО2, происходит таутомеризация присоединенной группы и образуется карбамат. Заметьте, что данная молекула СО2 не используется для карбокилирования – не включается в углеродные скелеты. Далее в активный центр последовательно заходят ион Mg2+ и RuBP. Mg2+ связывается с карбаматом и активирует его, создавая определенную активную конформацию. Функция карбамата заключается в катализе кето-енольной изомеризации RuBP – происходит депротонирование С3 на RuBP и образование нестабильной ендиольной формы (рисунок 6) с двойной связью между С2 и С3. Далее молекула воды атакует С3, а СО2 — С3 RuBP, разрывается связь между С2 и С3. Таким образом, получаются два трехуглеродных продукта реакции – два 3-фосфоглицерата.
Помимо описанного для активации RuBisCO необходимо взаимодействие с активазой RuBisCO – АТР-зависимым ферментом (т.е. тоже работает только на свету!). В темноте в активном сайте RuBisCO, не образовавшем карбамата, прочно удерживается RuBP, который блокирует работу фермента. Активаза RuBisCO высвобождает RuBP из активного сайта, делая его доступным для образования карбамата.
Более подробный механизм реакций, происходящих в активном сайте RuBisCO, можно посмотреть здесь: https://www.ebi.ac.uk/thornton-srv/m-csa/entry/907/.
На следующей стадии, стадии восстановления, происходят процессы, обратные реакциям гликолиза (рисунок 7): фосфорилирование 3-фосфоглицерата фосфоглицераткиназой до 1,3-дифосфоглицерата и затратой одной молекулы ATP, и дальнейшее восстановление 1,3-дифосфоглицерата ферментом глицеральдегидфосфатдегидрогеназой до глицеральдегид-3-фосфата с использованием в качестве донора электронов NADPH+H + и выделением свободного фосфата. Затем из глицеральдегид-3-фосфата в равновесной реакции, катализируемой триозофосфатизомеразой образуется дигидроксиацетонфосфат. Использование в реакциях стадии восстановления ATP и NADPH+H+ делает стадию сильно зависимой от энергии, а значит – и от солнечного света.
Образованные восстановленные триозы далее вступают в реакции стадии регенерации. На этой стадии происходят три основных типа реакций – альдолазная, транскетолазная и фосфатазная. Основные процессы схематично изображены на рисунке 8. Сначала в реакции, катализируемой ферментом альдолазой из глицеральдегид-3-фосфата и дигидроксиацетонфосфата образуется фруктозо-1,6-бисфосфат. Альдолазная реакция обратима, в ее механизме в активном центре фермента происходит образование протонированного Шиффового основания между С2 кетозы (дигидроксиацетонфосфата) и ε-аминогруппой остатка лизина, в ходе дальнейшей нуклеофильной реакции с С1 карбонильной группы альдозы (глицеральдегид-3-фосфата) образуется фруктозо-1,6-бисфосфат.
Далее фруктозо-1,6-бисфосфат гидролизуется до фруктозо-6-фосфата. Эту реакцию катализирует фруктозо-1,6-бисфосфатаза, и данная реакция необратима. Необратимость реакции позволяет сделать ее регуляторной для всего цикла.
Далее фруктозо-6-фосфат вступает с глицеральдегид-3 фосфатом в транскетолазную реакцию, в ходе которой на глицеральдегид-3-фосфат переносятся С1-С2 с фруктозо-6-фосфата. В данной реакции участвует кофермент тиаминпирофосфат (TPP), являющийся простетической группой транскетолазы. ТРР, связываясь углеродом тиазольного кольца с С2 атомом кетозы, вызывает расщепление ковалентной связи между С2 и С3 на кетозе и образование карбаниона с отрицательным зарядом на С2, к которому присоединяется С-атом альдегидной группы альдозы (глицеральдегид-3-фосфат), имеющий частичный положительный заряд. В ходе реакции образуются четырехуглеродный сахар эритрозо-4-фосфат и пентоза – ксилулозо-5-фосфат.
Образованный эритрозо-4-фосфат вступает в альдолазную реакцию с дигидроксиацетонфосфатом с образованием седогептулозо-1,7-бисфосфата, который затем гидролизуется до седогептулозо-7-фосфата. Реакция во многом похожа на гидролиз фруктозо-1,6-бисфосфата и так же является необратимой и регуляторной.
В следующей транскетолазной реакции седогептулозо-7-фосфат взаимодействует с глицеральдегид-3-фросфатом с образованием двух пентоз – рибозо-5-фосфата и ксилулозо-5-фосфата.
Итого, получены 3 различные пентозы. В последующих реакциях (рисунок 9) происходит эпимеризация ксилулозо-5-фосфата в рибулозо-5-фосфат под действием фермента рибулозофосфатэпимеразы и изомеризация рибозо-5-фосфата в рибулозо-5-фосфат под действием фермента рибозофосфатизомеразы. Полученные рибулозо-5-фосфаты фосфорилируются по С1 рибулозофосфаткиназой с образованием рибулозо-1,5-бисфосфата (RuBP). Данная реакция необратима. Таким образом, в ходе цикла Кальвина вновь образовался RuBP, который снова может вступать в реакцию карбоксилирования. Помимо этого был зафиксирован СО2.
Если сделать пересчет на три зафиксированных СО2, то есть – на одну синтезированную триозу, то суммарное уравнение ОПФП выглядит следующим образом:
3CO2 + 6NADPH + 6H+ + 9ATP → глицеральдегид-3-фосфат + 6NADP+ + 9ADP + 3H2O + 8Pi
Таким образом, для работы цикла Кальвина требуется соотношение ATP/NADPH+H+ = 1,5. Вспомним снова световую фазу, при нециклическом транспорте электронов на каждый восстановленный NADPH+H+ в люмене хлоропласта становится на 6 протонов больше (2Н+ на PSII, 2Н+ на цитохром-b6/f комплексе и 2H+ — в Q-цикле). АТФ-синтаза хлоропласт содержит 12-14 с-субъединиц, значит, для полного разворота F0 требуется 12-14 протонов и при этом образуется 3 ATP. Простые расчеты дают на этом основании стехиометрию H+/ATP = 4,7 (для 14 с-субъединиц) и ATP/NADPH+H+ = 1.3, что меньше необходимого, то есть ATP находится в недостатке. Для компенсации этого недостатка в ЭТЦ тилакоида идет циклический транспорт электронов вокруг I фотосистемы.
Кроме восстановительного пентозофосфатного пути существует еще и окислительный. Большинство ферментов и реакций этих путей общие и многие реакции являются обратимыми, ОПФП, как и ВПФП проходит в строме хлоропласта. Очевидно, что для нормального функционирования обоих путей и во избежание бессмысленной траты АТP необходимо регулировать активность ферментов, которая осуществляется с помощью тиоредоксиновой системы и изменения рН стромы хлоропласта (рис 10).
Помимо карбоксилазной активности RuBisCO проявляет также и оксигеназную (рисунок 11). В ходе этой реакции в активный центр RuBisCO вместо СО2 приходит О2 и образуются 2-фосфогликолат и только один 3-фосфоглицерат. Даже небольшие концентрации 2-фосфогликолата опасны для растения, вещество оказывает сильное и ингибирующее воздействие на многие системы и реакции. Для устранения последствий карбоксилазной активности RuBisCO в растениях существует целый метаболический путь, называемый фотодыханием, или гликолатный путь (не путать с глиоксилатным!), или С2-фотосинтез. Реакции данного пути требуют консолидированной работы сразу трех органелл – хлоропласта, пероксисомы и митохондрии (рисунок 12, из учебника Страсбургера «Физиология растений»).
Образованный в строме хлоропласта 2-фосфогликолат гидролизуется фосфогликолатфосфатазой и продукт, гликолат, выносится из хлоропласта через интегрированныйво внутреннюю мембрану транспортер и попадает в пероксисому через образованные поринами пор. В пероксисоме гликолат окисляется до глиоксилата гликолатоксидазой. Данная реакция идет в присутствии кислорода, на который через флафинмононуклеотид гликолатоксидазы передаются электроны и образуется H2O2. Н2О2 разлагается каталазой до воды и кислорода, именно поэтому данную реакцию необходимо проводить в пероксисоме. Далее ферментом глутамат-глиоксилатаминотрансферазой осуществляется переаминирование – перенос аминогруппы с глутамата на глиоксилат с образованием 2-оксоглутарата и глицина. В качестве донора аминогруппы может использоваться также аланин, помимо этого реакцию осуществляет и серин-глиоксилатаминотрансфераза (см. далее). Глицин затем поступает через аминокислотный транслокатор в митохондрию, где окисляется мультиферментным комплексом, сходным с пируватдегидрогеназным комплексом цикла Кребса, полное название которого – глициндекарбоксилазосерингидроксиметилтрансферазный комплекс (рисунок 13). В ходе реакций, катализируемых данным комплексом глицин, связываясь с пиридоксалем, образует основание Шиффа (B) и декарбоксилируется, затем остаток глицина передается на липоевую кислоту (C) и дезаминируется, окисляясь до формильного остатка, который связывается с тетрагидрофолатом (D, 1, 2). Восстановленная при этом дигидролипоевая кислота окисляется флафинадениндинуклеотидом (FAD) снова до липоевой кислоты, при этом FADH2 восстанавливает NADP+ (D-E). Формильный остаток с тетрагидрофолата передается на вторую молекулу глицина, образуя серин (А-В).
Таким образом, в ходе данной реакции из двух молекул глицина образуется серин, аммоний, CO2 и NADH+H+. Серин затем транспортируется из митохондрии обратно в пероксисому, гда происходит его дезаминирование серин-глиоксилатаминотрансферазой с образованием гидроксипирувата. Фермент гидроксипируватредуктаза восстанавливает гидроксипируват до глицерата, донором электронов для реакции является NADH+H+, глицерат транспортируется гликолат-глицератным переносчиком в строму хлоропласта и там фосфорилируется глицераткиназой. Итого, имеем регенерированный из 2-фосфогликолата 3-фосфоглицерат.
Аммоний, образованный в ходе дыхания, также переносится в хлоропласт, где с использованием ATP ферментом глутаминсинтетазой осуществляется аминирование глутамита до глутамина, а затем глутаматоксоглутаратаминотрансфераза (ГОГАТ) из глутамина и 2-оксоглутарата при использовании двух восстановленных ферредоксинов (то есть реакция фиксации аммония в листьях тоже светозависима!) в качестве доноров электронов катализирует образование двух глутаматов.
Полученный при декарбоксилировании глицина СО2 может быть снова зафиксирован RuBisCO. Можно привести некоторый подсчет «стоимости» оксигеназной активности RuBisCO. На две пентозы при связывании RuBisCO двух молекул кислорода происходит образование двух 3-фосфоглицератов и двух 2-фосфогликолатов. По стехиометрии фотодыхания на два 2-фосфогликолата образуется один 3-фосфогликолат (два синтезированных глицина превращаются в один серин), при этом тратится 1ATP на фосфорильрование глицерата и 1 АTP + NADPH+H+ (энергетический эквивалент двух ферредоксинов) – при фиксации аммония. При рефиксации выделившегося CO2 в ВПФП образуется 1/3 3-фосфоглицерата с использованием 3 АТР и 2 NADPH+H+ (смотри стехиометрию пути). То есть, из расчета на два акта оксигеназной активности можно получить 2+1+0,33 = 3,33 3-фосфоглицерата. Далее при синтезе из них в ВПФП двух RuBP (учтя, что 3C*3,(3) = 2*5C) используются еще пять ATP и три NADPH+H+. Итого выходит для АТР: 1+1+3+5=10; для NADPH+H+: 1+2+3=6.

Для просмотра таблицы прокрутите страницу вправо
Для просмотра таблицы прокрутите страницу вправо
|
Затраты на соответствующую реакцию на 1 моль газа |
Затраты (моль) |
|
|
ATP |
NADPH+H+ |
|
|
Карбоксилазная |
3 |
2 |
|
Оксигеназная |
5 |
3 |
Состояние, когда весь фиксируемый CO2 расходуется из-за оксигеназной активности, получило название углекислотной компенсации. Нетрудно догадаться из стехиометрии ВПФП и фотодыхания, что такая ситуация будет наблюдаться при соотношении карбоксилазной и оксигеназной реакций = 1:2.
Возникает резонный вопрос – почему миллиарды лет эволюции не позволили RuBisCO избавиться от оксигеназной активности или существенно ее снизить. Одно из объяснений заключается в том, что RuBisCO, как ферментный комплекс появился задолго (по разным данным — более миллиарда лет) до кислородной катастрофы и эволюционировал в условиях, когда оксигеназная активность не мешала в силу отсутствия субстрата. Несмотря на появление оксигенного фотосинтеза порядка 2,8-2,4 млрд лет назад, кислород в атмосфере стал накапливаться и достигать современного уровня лишь в последние пять-шесть сотен миллионов лет. Возможно, сложность комплекса не позволила ему адаптироваться и уменьшить уровень оксигеназной активности. Другое, более «физиологичное» объяснение наличия у RuBisCO оксигеназной активности – адаптация к аридным местам обитания с избыточной инсоляцией. Живущие в таких условиях растения вынуждены избавляться от избытка энергии и перевосстановленности, образующихся в ходе световых реакций и приводящих к повреждениям клетки. Использовать эту избыточную энергию на фиксацию СО2 невозможно, так как закрыты устьица, чтобы не допустить потерю воды. Поэтому в данном случае затратное фотодыхание служит защитой фотосинтетического аппарата.
Рассмотренные процессы происходят в растениях с так называемым С3 фотосинтезом, по количеству углеродов в первом образованном после фиксации СО2 веществе – 3-фосфоглицерате. Но у растений существуют и другие типы фотосинтеза, их много, если учесть переходные формы. Далее мы рассмотрим некоторые основные.
Концентрация кислорода в современной атмосфере равна 21%, СО2 – порядка 0,035-0,04%. Известно, что с повышением температуры растворимость газов уменьшается, а так как СО2 всего лишь около 350 ppm, в водном растворе СО2 становится очень мало по сравнению с содержанием О2, падает соотношение CO2/O2, что способствует значительному возрастанию оксигеназной активности. Растения научились бороться с этим и выживать в засушливых условиях с повышенной температурой и инсоляцией. Ключевая реакция здесь – первичная фиксация углекислого газа ферментом фосфоенолпируваткарбоксилазой (ФЕП-карбоксилаза, PEPCase).
В одном случае CO2 в форме карбоксильной группы низкомолекулярных органических веществ (малат, оксалоацетат, аспартат) перемещается из клетки первичной фиксации (клетка хлоренхимы мезофилла) в клетку обкладки пучка (рис. 14), где происходит декарбоксилирование и образование СО2. Тут СО2 уже фиксируется с помощью RuBisCO. Данный механизм позволяет создать в области активности RuBisCO условия с повышенной концентрацией СО2, благодаря чему можно если не убрать, то минимизировать оксигеназную реакцию. Схематичный процесс, который называется в честь первооткрывателей циклом Хэтча-Слэка-Карпилова (хотя последнего все забывают), представлен на рисунке 14. Такой фотосинтез называется С4 из-за того, что первоначально при первичной фиксации образуется четырехуглеродное соединение – оксалоацетат.
Листья растений с С4 типом фотосинтеза имеют своеобразную анатомию (рисунок 15), называемую кранц-анатомией (нем. Kranz – венок). Кранц-анатомия и С4 фотосинтез эволюционно возникала независимо во многих таксонах цветковых растений и имеет различное строение в зависимости от типа С4 фотосинтеза и видовой принадлежности самого растения, более того, даже помимо переходных случаев, вроде С3-С4 фотосинтеза, есть данные, что у типичных С3 растений вполне может проходить и С4 фотосинтез.
ФЕП-карбоксилаза катализирует образование оксалоацетата из иона бикарбоната и фосфоенолпирувата. На первой стадии реакции происходит нуклеофильная атака бикарбонатом фосфата фосфоенолпирувата, образуются два промежуточных продукта – фосфокарбонат и енольная форма пирувата. На второй, необратимой стадии, происходит разрыв эфирной связи в фосфокарбонате, выходит из реакции Pi, а CO2 реагирует с енольной формой пирувата с образованием оксалоацетата. У С4 растений ФЕП-карбоксилаза активируется фосфорилированием осуществляемым ферментом киназой ФЕП-карбоксилазы, при этом сама киназа активируется светом, в темноте ФЕП карбоксилаза инактивируется фосфорилазой, гидролизующей присоединенный на свету фосфат. Поэтому данная стадия у С4 растений идет только на свету. У ФЕП-карбоксилазной реакции два больших преимущества: (1) отсутствие оксигеназной активности и (2) использование бикарбоната в качестве
субстрата. Образование HCO3— из СО2 (помимо самопроизвольного) катализируется карбоангидразой. Данный факт позволяет накопить большое количество углерода в форме HCO3-, ведь в равновесии с 8 мкмоль СО2 при 25°С и рН=8 находится 400 мкмоль HCO3-, таким образом, минимизируется проблема малого количества растворенного СО2 при высокой температуре.
Так как в С4 фотосинтезе разные клетки и разные органеллы объединены в один метаболический процесс, необходима система транспортеров для различных переносимых соединений. Например, малат-оксалоацетатный и малат-аспартатный челноки. Во внутренних мембранах митохондрий и пластид располагается транслокатор, обменивающий, например, малат на оксалоацетат и наоборот. Таким образом, данные соединения могут находиться в строме хлоропласта, матриксе митохондрии, выходить оттуда в цитоплазму, по градиенту диффузионного потенциала перемещаться по симпласту между клетками и снова заходить в матрикс митохондрии или строму хлоропласта. Помимо этого, благодаря ферменту малатдегидрогеназе, который осуществляет окисление малата с образованием оксалоацетата и восстановлением NAD(P)H+H+ (существуют изозимы, специфичные как к NAD+, так и к NADP+), а также и обратную реакцию, через мембрану транспортируется и восстановительный эквивалент. Также важным является триозофосфат-3-фосфоглицератный челнок (обмен дигидроксиацетонфосфата и 3-фосфоглицерата), который, помимо восстановительного эквивалента, переносит еще и энергию, которая конвертируется в АТР (рис. 20).
Есть и большое количество других переносчиков, обменивающих аминокислоты, фосфаты, нуклеотиды, ионы и проч.
С4 тип фотосинтеза можно разделить на три основных подтипа (см. рисунки 19,21,22) – NADP+-ME, NAD+-ME и ФЕП-карбоксикиназный. Все они названы по ферменту, который осуществляет декарбоксилирование, соответственно – NADP+-зависимая декарбоксилирующая малатдегидрогеназа или NADP+ -малик-энзим, NAD+ -малик-энзим и ФЕП-карбоксикиназа. Ниже рассмотрим подробнее каждый из типов.
Самым простым, пожалуй, является NADP+-ME тип. Пируват в клетках мезофилла транспортируется в хлоропласт и там в реакции, катализируемой пируватортофосфатдикиназой (PPDK) из него образуется фосфоенолпируват (ФЕП). Субстратами PPDK являются АТР, фосфат и пируват, продуктами реакции – пирофосфат и AMP. Пирофосфат в строме быстро гидролизуется пирофосфатазой до двух фосфатов, что сильно сдвигает реакцию образования ФЕП в сторону продуктов. PPDK активна только на свету, в темноте происходит ее фосфорилирование, причем в данном случае используется не АТР, а ADP, которого как раз много в темноте, а не на свету.
ФЕП далее поступает из хлоропласта в цитоплазму, где происходит карбоксилазная реакция, катализируемая ФЕП-карбоксилазой, которая была описана выше. Образованный в ходе реакции оксалоацетат снова поступает в хлоропласт, где происходит его восстановление до малата NADPH+-зависимой малатдегидрогеназой. Малат выходит из хлоропласта в цитоплазму и по симпласту через плазмодесмы по градиенту концентрации (в зону меньшей концентрации) переносится в цитоплазму клетки обкладки. В клетках обкладки малат поступает в строму хлоропласта, здесь происходит реакция декарбоксилирования ферментом NADP+ -малик-энзимом с образованием пирувата, углекислого газа и восстановленного NADPH+H+. Пируват далее снова выходит из хлоропласта и диффундирует в клетку мезофилла, СО2 используется в карбоксилазной реакции RuBisCO, NADPH+H+ в цикле Кальвина.
У растений с NADP+-ME типом фотосинтеза клетки мезофилла имеют мощно развитую гранальную структуру пластид, а пластиды клеток обкладки, наоборот, агранальные. Возникает проблема – как в агранальных хлоропластах обеспечить энергией и восстановительными эквивалентами протекающий цикл Кальвина. Для решения этой проблемы и испольхуется упомянутый выше триозофосфат-3-фосфоглицератный челнок (рис. 20).
В NAD+-ME типе (рис. 21) в клетках мезофилла происходит переаминирование, катализируемое аспартатаминотрансферазой, полученный в ходе реакции из оксалоацетата аспартат диффундирует в клетку обкладки и там поступает уже не в строму хлоропласта, а в матрикс митохондрии. В митохондрии происходит обратная реакция и снова образуется оксалоацетат, который, как и в предыдущем случае NAD(P)+-зависимой малатдегидрогеназой восстанавливается до малата. Малат декарбоксилируется до пирувата с выделением СО2 и восстановлением NAD+ NAD+-малик-энзимом. Углекислый газ диффундирует в хлоропласт клетки обкладки. Пируват выходит из стромы митохондрии в цитоплазму, и там происходит его аминирование аланинаминотрансферазой. Аланин диффундирует в клетку мезофилла, где снова дезаминируется и превращается в пируват.
Хлоропласты клеток обкладки в растениях с В NAD+-ME типом фотосинтеза имеют гранальную структуру, в то время как клеток мезофилла – наоборот, их структура может быть агранальной.
ФЕП-карбоксикиназный вариант С4 фотосинтеза более сложный. Образованный в ФЕП-карбоксилазной реакции оксалоацетат имеет разную дальнейшую судьбу. Он может, как и в случае NAD+-ME фотосинтеза, преобразуясь в аспартат, поступать в клетки обкладки пучка, а может, как в NADP+-ME типе, поступать снова в хлоропласт мезофилла, превращаться в малат и уже в малат будет диффундировать в клетку обкладки (данная часть на рисунке 22 не показана). Малат далее поступает в митохондрию клетки обкладки, где декарбоксилируется, и дальнейшие реакции повторяют реакции уже NAD+-ME типа фотосинтеза. В том случае, если оксалоацетат превращается в аспартат, аспартат диффундирует в цитоплазму клеток обкладки там дезаминируется аспартатаминтрансферазой, и оксалоацетат вступает в реакцию, катализируемую ФЕП-карбоксикиназой. В ходе этой реакции с использованием АТР оксалоацетат декарбоксилируется и превращается в ФЕП. ФЕП дальше диффундирует обратно в клетки мезофилла, где может снова вступить в карбоксилазную реакцию. Таким образом, в данном типе С4 фотосинтеза сосуществуют две декарбоксилирующие реакции, производящие СО2 для RuBisCO.
С4 тип фотосинтеза происходит в основном у растений, обитающих в климате с повышенной температурой и отчасти с недостатком воды (рис 23). Если брать в рассмотрение только протекающие в процессе фиксации углерода реакции, то можно прийти к выводу, что по сравнению с C3 фотосинтезом С4 довольно расточителен. Действительно, в условиях умеренного климата с относительно стабильным и достаточным увлажнением растения с С3 фотосинтезом имеют преимущество перед С4 растениями, опережая их в скорости накопления биомассы. Но как только растения оказываются в жарком климате, энергетические затраты на дополнительные реакции у С4 растений м перекрываются большим выигрышем в уровне фиксации СО2 и уменьшении оксигеназной активности RuBisCO.
Растения, произрастающие в засушливых с долговременной нехваткой влаги или даже полным отсутствием воды в течение продолжительного времени, развили третий тип фотосинтеза. CAM-фотосинтез (от англ. Crassulacean acid metabolism – метаболизм по типу толстянковых) довольно широко распространен среди высших растений, хоть и был впервые описан на семействе Crassulaceae. САМ фотосинтез, точно так же, как и С4, возникал у разных групп растений неоднократно и независимо друг от друга. Растения с САМ фотосинтезом встречаются и во многих группах – представителей можно встретить среди плауновидных, папоротникообразных, гнетовых, голосеменных и покрытосеменных растений. Точно так же есть растения с переходными типами фотосинтеза, существует много примеров изменения типа фотосинтеза с С3 на САМ у одного растения в зависимости от внешних условий и/или стадии онтогенеза и даже от рассматриваемого органа. САМ фотосинтез в основном выполняет роль физиологической адаптации к недостатку воды, потому встречается не только у пустынных растений, но так же у эпифитов и галофитов. Помимо этого, САМ фотосинтез имеет распространение и среди водных растений (например, Isoetes howellii или Crassula aquatica). Недостатка воды в данном случае растение не испытывает, но сталкивается с недостатком СО2 в силу его низкой концентрации и малой доступности, особенно в кислых водоемах, где равновесие сдвинуто в сторону образования СО2 из растворенного бикарбоната. Такие растения фиксируют СО2 ночью, когда не так высока конкуренция за СО2 с другими фотосинтетиками, населяющими водоем, а также когда эти фотосинтетики не образуют кислород.
САМ фотосинтез с точки зрения биохимии процессов очень похож на С4, основная его особенность в том, что происходит разделение первичной и вторичной фиксации СО2 не в пространстве, а во времени. В данном случае первичная фиксация с помощью ФЕП-карбоксилазы происходит ночью, в вечерние и ранние утренние часы – то есть в то время, когда у растений открыты устьица, а активность RuBisCO наблюдается днем, когда устьица закрыты, чтобы максимально уменьшить испарение воды.
В отличие от С4 типа фотосинтеза, в данном случае ФЕП-карбоксилаза работает не на свету, а ночью и светом наоборот ингибируется. Работа данного варианта фермента контролируется циркадными ритмами на уровне транскрипции киназы ФЕП-карбоксилазы – транскрипты этого фермента накапливаются именно в темное время суток.
Начнем рассмотрение САМ фотосинтеза с процессов, происходящих ночью. В темное время суток крахмал, который накопился в ходе протекания цикла Кальвина на свету, гидролизуется под действием различных ферментов (фосфорилаза и разные амилазы), образованные моно- и дисахариды превращаются в глюкозо-6-фосфат, который затем превращается в триозофосфат (реакции, аналогичные начальным стадиям гликолиза). Триозофосфаты выходят в цитоплазму, где протекают последующие реакции гликолиза, завершающиеся формированием фосфоенолпирувата. ФЕП вступает в реакцию с гидрокарбонатом, катализируюмую ФЕП-карбоксилазой и затем образованный оксалоацетат восстанавливается до малата. Малат (в виде аниона) закачивается в выкуоль. В вакуоль с помощью вакуолярной протонной помпы (ATPase V-типа) закачиваются противоионы – H+. Таким образом, рН вакуоли за ночь очень сильно понижается (вплоть до рН=3), этим обусловлен кислый вкус САМ растений в предрассветные и утренние часы. Энергетически затратная закачка протонов через тонопласт необходима для уменьшения осмотического давления, которое могло бы быть создано малатом калия. Яблочная кислота с обеими протонированными карбоксильными группами создает в три раза меньшее осмотическое давление, чем калиевая соль. Во время всех описанных событий устьица растений остаются открытыми и СО2 свободно диффундирует в межклетники.
Утром на свету ФЕП-карбоксилаза довольно быстро ингибируется, устьица закрываются. При этом малат выходит из вакуоли и декарбоксилируется. Эту реакцию могут осуществлять, в зависимости от конкретного растения, разные декарбоксилирующие ферменты — различные малик-энзимы или ФЕП-карбоксикиназа, декарбоксилирующая синтезированный из малата оксалоацетат. Соответственно типу декарбоксилирующегофермента, дальнейшие процессы происходят либо в митохондриях, либо в хлоропластах. Так или иначе, высвободившийся СО2 поступает в хлоропласт и там связывается RuBisCO. Образованный в ходе цикла Кальвина триозофосфат либо вступает в путь глюконеогенеза и образует в хлоропласте пул крахмала, который будет дальше гидролизоваться ночью для синтеза малата, либо поступает в цитоплазму, где из него синтезируется сахароза, которая отправляется в дальний транспорт по растению.
biocpm.ru
Темновая фаза фотосинтеза
Продукты световой фазы фотосинтеза (АТФ и НАДФН) используются в темновой фазе для восстановления СО2 до уровня углеводов. Реакции восстановления происходят настолько быстро, что с помощью обычных методов химического анализа чрезвычайно трудно обнаружить промежуточные продукты и практически невозможно установить последовательность их превращений. Только использование радиоактивных изотопов, в первую очередь 14С, хроматографии и электрофореза позволило установить последовательность реакций при фотосинтезе, множественность путей метаболизма углерода. В настоящее время известны С3— и С4-пути фиксации СО2, фотосинтез по типу толстянковых (САМ-метаболизм) и фотодыхание.
С3-путь фотосинтеза (цикл кальвина)
Этот способ ассимиляции СО2, присущий всем растениям, был расшифрован американскими исследователями во главе с Мэльвином Кальвиным. Схема последовательности реакций получила название цикла Кальвина. Характерной особенностью фотосинтетического восстановления СО2 являются цикличность и разветвленность этого процесса. Цикличность обеспечивает высокую производительность, саморегуляцию и непрерывность образования углеводов. Разветвленность — образование разнообразных продуктов, дублирование путей регенерации акцептора СО2. Цикл состоит из трех этапов: карбоксилирования, восстановления и регенерации акцептора СО2 (рис.).
Карбоксилирование. Первой реакцией, вводящей СО2 в цикл Кальвина, является карбоксилирование рибулезо-1.5-дифосфата (1,5-РДФ) с участием фермента рибулезодифосфаткарбоксилазы (РДФ-карбоксилазы). Образующееся при этом нестойкое щестиутлеродное соединение быстро распадается на триозы — две молекулы 3-фосфоглицериновой кислоты (З-ФГК). Поэтому З-ФГК можно считать первичным продуктом фотосинтеза.
Фаза восстановления. Восстановление 3-фосфоглицериновой кислоты (З-ФГК) до 3-фосфоглицеринового альдегида (3-ФГА) происходит в два этапа. Сначала при участии АТФ и фосфоглицераткиназы З-ФГК присоединяет остаток фосфорной кислоты с образованием 1,3-дифосфоглицериновой кислоты (1,3-ДФГК.) Этим достигается повышение реакционной способности соединения и возможность его восстановления с помощью НАДФ-Н до З-ФГА. Это единственная восстановительная реакция цикла. Фаза восстановления является центральным звеном цикла. Именно здесь скрещиваются световая и темновая фазы фотосинтеза. Все остальные превращения идут на уровне сахаров, одинаковых по степени восстановленности.
Фаза регенерации первичного акцептора СО2 и синтеза конечных продуктов фотосинтеза. В результате рассмотренных ранее реакций при фиксации трех молекул СО2 образуются 6 молекул восстановленных 3-фосфотриоз, пять из них используются затем для регенерации РДФ, а 1- для синтеза глюкозы. Это достигается следующей последовательностью реакций. Часть молекул З-ФГА под действиеч триозофосфатизомеразы превращается в фосфодиоксиацетон (ФДА). Затем два изомера (ФГА и ФДА) полвергаются конденсации при участии фермента альдолазы, образуя фруктоза-1,6-дифосфат, у которого затем отщепляется один фосфат.
В дальнейших реакциях, связанных с регенерацией акцептора СО2, возникает цепь фосфорных эфиров сахаров, содержащих в своем составе 4, 5 или 7 атомов углерода. В этих реакциях последовательно принимают участие транскетолазы и трансальдолазы. Транскетолаза катализирует перенос 2-углеродной группировки от фруктоза-6-фосфата на З-ФГА, при этом образуются эритрозо-4-фосфат (С4) и ксилулозо-5-фосфат (С5). Затем альдолаза осуществляет перенос 3-углеродного остатка ФДА на эритрозо-4-фосфат, в результате чего синтезируется седогептулозо-1.7-дифосфат (С7). От последнего отщепляется один остаток фосфорной кислоты и под действием транскетолазы из него и З-ФГА образуются ксилулозо-5-фосфат (С5) и рибозо-5-фосфат (С5). Две молекулы ксилулозо-5-фосфата путем эпимеризации и одна молекула рибозо-5-фосфата за счет изомеризации превращаются в три молекулы рибулозо-5-фосфата (С5). Это соединение подвергается фосфорилированию за счет АТФ, образующейся в световую фазу.
Из оставшейся неиспользованной молекулы ФГА при повторении цикла образуется фруктоза-1,6-дифосфат, из которой могут образовываться глюкоза, сахароза и крахмал.Таким образом, для синтеза одной молекулы ГЛЮКОЗЫ (С6) должно произойти шесть оборотов цикла. В каждом обороте используются 3 мол. АТФ (две для активирования двух молекул ФГК и одна при регенерации акцептора СО2 1,5-РДФ) и две молекулы НАДФН для восстановления ФГК до ФГА. Поэтому для синтеза одной молекулы глюкозы в цикле Кальвина необходимо 12 НАДФН и 18 АТФ. Таков энергетический вклад световой фазы в темновую фиксацию СО2.
В цикле Кальвина первичными продуктами включення СО2 в органические вещества н восстановления являются трехуглеродные соединения (З-ФГК, З-ФГА, ФДА). Поэтому этот способ фиксации СО2 носит название С3-пути фотосинтеза. Большинство растений, особенно произрастающих в умеренной зоне, используют именно этот путь восстановления СО2 и называются С3-растениями.
studfile.net
4. Световая фаза фотосинтеза
Во время световой фазы фотосинтеза происходят стабилизация и запасание световой энергии и ее трансформация в химическую энергию. Обязательным условием для осуществления световой стадии фотосинтеза является наличие света.
Световая фаза фотосинтеза включает 4 основных процесса:
1) фотохимическое возбуждение хлорофилла;
2) фотоокисление (фотолиз) воды до кислорода, протонов и электронов;
3) фотовосстановление НАДФ окисленного до НАДФ восстановленного;
4) фотосинтетическое фосфорилирование (образование АТФ из АДФ и фосфорной кислоты при участии энергии света).
Внутренняя мембрана хлоропластов непроницаема для протонов.
Реакционный центр ФС 1 под действием кванта света переходит в возбужденное состояние, он окисляется и отдает электроны на ферредоксин, который передает их на фермент редуктазу. Редуктаза окисляет НАДФ+ до НАДФ Н. Под действием кванта света реакционный центр ФС 1 переходит в возбужденное состояние и окисляется, отдавая 2 электрона на комплекс переносчика.
Таким образом, в полости тилакоида накапливаются протоны, в результате возникает разность концентраций протонов между сторонами мембраны и разность зарядов.
За счет энергии электрохимического потенциала конформация фермента АТФазы изменяется, канал открывается и по нему протоны возвращаются в строму. Одновременно в головке АТФ-азы из АДФ и неорганического фосфата синтезируется АТФ.
5. Темновая фаза фотосинтеза
Темновая фаза фотосинтез – это путь восстановления двуокиси углерода до простых сахаров. Этот процесс впервые был изучен американским ученым Кальвином, поэтому в его честь был назван циклом Кальвина.
Цикл Кальвина функционирует у высших растений, водорослей, у большинства фототрофных бактерий.
Первичным акцептором углекислого газа является рибулозо-1,5-бисфосфат. Ключевым ферментом, обеспечивающим фиксацию СО2, является фермент рибулозобисфосфаткарбоксилазаоксигеназа (Рубиско). Его активность строго коррелирует с общей интенсивностью фотосинтеза. Концентрация его в строме очень высока. На долю этого фермента приходится почти 60% растворимого белка стромы.
Цикл Кальвина, весьма напоминающий обращенный пентозофосфатный путь дыхания, состоит из 4-х этапов:
Стадия карбоксилирования: молекулы рибулозо-5-фосфата фосфорилируются с участием АТФ и фермента фосфорибулокиназы, в результате чего образуются молекулы рибулозо-1,5-бисфосфата. К ним, в свою очередь, присоединяется углекислый газ с помощью фермента Рубиско. Полученный продукт расщепляется на 2 триозы – 2 молекулы 3-ФГК.
Стадия восстановления ФГК. Образовавшиеся на предыдущем этапе молекулы 3-ФГК восстанавливаются до альдегида 3-ФГА в 2 этапа. Сначала происходит фосфорилирование 3-ФГК при участии АТФ и фосфоглицераткиназы до 1,3-бисфосфоглицериновой кислоты, а затем ее восстановление до фосфоглицеринового альдегида (при участии НАДФ восстановленного).
3) Стадия регенерации первичного акцептора СО2. Происходит с участием фосфатов трех-, четырех-, пяти-, шести- и семиуглеродных сахаров. В результате процессов взаимопревращений фосфорилированных сахаров из 5 молекул трехуглеродных сахаров образуются 3 молекулы пятиуглеродных сахаров, которые затем превращаются в рибулозобисфосфат.
4) Стадия синтеза углеводных продуктов фотосинтеза. На этом этапе из фосфоглицеринового альдегида и фосфодиоксиацетона под действием фермента альдолазы синтезируется фруктозо-1,6-бисфосфат, который затем может превращаться или в крахмал, или сахарозу.
Для синтеза 1 молекулы глюкозы в цикле Кальвина необходимо 12 молекул НАДФ восстановленного и 18 молекул АТФ.
Процесс фотосинтеза осуществляется только на свету. Однако у растений часто возникает потребность в глюкозе, когда фотосинтез невозможен (в темное время суток, при прорастании семян). В этих случаях потребности в глюкозе удовлетворяются либо за счет распада сложных углеводов, либо путем новообразования глюкозы, которое называется глюконеогенезом.
Глюконеогенез – процесс образования глюкозы из неуглеводных предшественников: пирувата, аминокислот, глицерина, жирных кислот и др.). По своему химизму этот процесс напоминает обращенный гликолиз.
Глюконеогенез происходит и в организмах человека и животных. Однако ферменты, которые катализируют реакции гликолиза, имеются только в клетках печени и надпочечников. Поэтому глюконеогенез может осуществляться только в этих органах, он идет в тех случаях, когда возникает потребность в глюкозе (например, у клеток головного мозга), а запасов гликогена недостаточно и другие возможности обеспечить эти потребности отсутствуют.
studfile.net
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит активирование молекул хлорофилла за счет энергии солнечного света. Избыточная энергия молекул хлорофилла используется на синтез АТФ и фотолиз (расщепление) воды на ионы Н+и ОН—. Эти процессы носят ферментативный характер и происходят с участием системы переносчиков электронов. Атомы водорода участвуют в восстановлении НАДФ, а ионы ОН—, взаимодействуя друг с другом, образуют молекулярный кислород и воду. Молекулярный кислород выделяется в атмосферу, а АТФ и НАДФ.Н2используются для восстановления углерода в темновой фазе фотосинтеза.
Темновая фаза фотосинтеза
Темновая фаза фотосинтеза включает реакции, идущие за счет энергии, накопленной в световую фазу или при дыхании. В темновой фазе фотосинтеза выделяют несколько этапов:фиксация СО2рибулозобифосфатом;
перенос активного водорода на соединение, в котором фиксирована поглощенная молекула СО2;
восстановление акцептора СО2;
образование сахаров.
(Подробнее о процессе фотосинтеза см. раздел «Общая биология»).
Дыхание
Дыхание — одна из важнейших сторон обмена веществ. Оно служит источником энергии, необходимой для осуществления всех процессов жизнедеятельности растения.К

Рис. 30. Дыхание листа.
ак и любой орган, лист интенсивно дышит, то есть поглощает кислород и выделяет углекислый газ. Причем процесс дыхания осуществляется постоянно, как на свету, так и в темноте. Если поместить в сосуд свежесрезанные листья, плотно закрыть его и поставить в темное теплое место, то на следующий день можно обнаружить, что состав воздуха в сосуде изменился. Если поместить в него свечу, то она погаснет, а известковая вода мутнеет. Этот простой опыт доказывает, что в сосудеуменьшилось количество кислорода и увеличилось количество углекислого газа, то есть листья дышат (рис. 30).
В отличие от фотосинтеза, во время дыхания происходит освобождение энергии, органические вещества окисляются, и происходит выделение углекислого газа.
Дыхание представляет собой цепь окислительно-восстановительных реакций, катализируемых ферментами. Эти реакции протекают в митохондриях клеток. (Механизм клеточного дыхания см. раздел «Общая биология»).
Транспирация
Под транспирациейпонимают испарение воды листьями (рис. 31). Транспирация обеспечивается ря-д
Рис. 31. Испарение воды листьями.
ом анатомических и физиологических механизмов и поэтому является физиологическим процессом, в отличие от чисто физического испарения воды.Значение транспирации очень велико, так как обеспечивает
терморегуляцию растения;
деятельность верхнего концевого двигателя водного тока.
Различают два вида транспирации — кутикулярную и устьичную.
Кутикулярная транспирация
Кутикулярная транспирацияпредставляет собой процесс испарения воды с поверхности кутикулы, покрывающей лист. Кутикулярная транспирация составляет порядка 10-20% общего испарения воды взрослыми листьями.Устьичная транспирация
Под устьичной транспирациейпонимают процесс испарения воды листьями с помощью особых структур — устьиц. Устьичная транспирация является основным механизмом водообмена между растением и атмосферой.Процесс устьичной транспирации можно разделить на 3 основные этапа:
испарение воды с поверхности клеток в межклетники;
выход паров воды из межклетников через устьичную щель;
диффузия паров воды от поверхности листа в более далекие слои атмосферы.
В настоящее время общепризнана гипотеза устьичных движений, связанная с перераспределением ионов калия между замыкающими и сопутствующими клетками, приводящим к перемещению воды. Причем основную роль в создании осмотического давления в замыкающих клетках играют ионы калия, которые закачиваются в них, и образующаяся на свету глюкоза.
Определенную роль играет и концентрация СО2. Избыток СО2, по-видимому, вызывает подкисление цитоплазмы. Это приводит к изменению рН, что приводит к закрыванию устьиц.
Таким образом, устьица чутко реагируют на изменения внешних условий и физиологические изменения в тканях листа, что влияет на интенсивность транспирации, приспосабливая ее к конкретным условиям.
При открывании устьиц углекислота поступает в лист и создает предпосылки для фотосинтеза. В отсутствие света фотосинтез в замыкающих клетках прекращается (как и во всех других), тургорное давление снижается и устьица закрываются. При недостатке поступления воды в растение устьица тоже закрываются, сберегая таким образом то небольшое количество влаги, которое доступно растению.
studfile.net
Темновая фаза фотосинтеза. Хемосинтез
При фотосинтезе темновая фаза следует за световой и неразрывно с ней связана. В ходе нее синтезируется глюкоза из углекислого газа. При этом затрачивается энергия АТФ и НАДФН. Разберем в деталях, что же происходит в темновой фазе.
Цикл Кальвина
1. Из рибулозомонофосфата при расходовании АТФ создается рибулозодифосфат — первичный акцептор СО2 (на 6 молекул рибулозомонофосфата расходуется 6 молекул АТФ). Считается, что рибулозодифосфата всегда достаточно в строме.
2. 1,5 рибулозодифосфат (дважды фосфорилированное производное рибозы) соединяется с СО2 с помощью фермента 1,5 дифосфаткарбоксилазы (реакция карбоксилирования). В результате образуется короткоживущее шестиуглеродное промежуточное соединение (карбоксирибитолфосфат, 6 молекул), которое вследствие гидролиза распадается на 2 молекулы фосфоглицериновой кислоты (фосфоглицерат). Так как для синтеза 1 молекулы глюкозы необходимо 6 молекул СО2, количество молекул кислоты умножаем на 6 и получаем 12 молекул фосфоглицериновой кислоты.
3. Еще 12 молекул АТФ затрачивается на синтез 12 молекул дифосфоглицериновой кислоты (дифосфоглицерат). Процесс катализируется ферментом фосфоглицераткиназой.
4. Затем под влиянием другого фермента (глицеральдегид-3-фосфат-дегидрогеназа) при окислении 12 молекул НАДФH образуется 12 молекул триозофосфата (фосфоглицеринового альдегида — ФГА, другое название — глицеральдегид-3-фосфат). Куда пойдут эти 12 молекул? Из 2 молекул триозофосфата (они выпадут из цикла) образуется фруктозо-6-фосфат, а из него 1 молекула глюкозы.
5. Другая часть триозофосфата (10 молекул) продолжает цикл, участвуя в превращении углеродных соединений, и преобразуется в рибулозомонофосфат (6 молекул). Цикл замыкается.
В опытах хлоропласты изолировали, помещали в темноту, чтобы выяснить, может ли в этом случае идти темновая фаза фотосинтеза. Она проходила только тогда, когда в хлоропласты добавляли АТФ и НАДФH — их необходимо было получить из световой фазы для нормального протекания темновой. В этом и состоит значение световой фазы.
Хемосинтез
1. Что позволяет бактериям без хлорофилла создавать органические вещества? Энергия, выделяющаяся в результате химических реакций окисления целого ряда разных неорганических соединений: аммиака, сероводорода, водорода, оксида железа (II) и других. Полученная энергия расходовалась на самостоятельный синтез органических веществ.
2. Следовательно, хемосинтез — это автотрофный тип питания. Он был открыт в 1889—1890 годах крупным микробиологом С. Н. Виноградским.
3. Серобактерии живут в водоемах, например, ими богато Черное море. Каковы химические основы их существования? Бактерии окисляют сероводород, а серу копят в себе: 2H2S + 02 = 2H20 + 2S + Е. Могут серобактерии окислять и серу до серной кислоты: 2S + 302 + 2H20 = 2H2S04+ Е.
4. Куда идет энергия, образованная в результате процесса окисления? На синтез органических веществ. При фотосинтезе энергия вырабатывается из солнечного света — в этом и состоит отличие фотосинтеза от хемосинтеза. Но потом она расходуется в том числе и на синтез органики — в этом сходство фотосинтеза с хемосинтезом.
5. Как действуют нитрифицирующие бактерии? Во-первых, идет окисление аммиака бактериями нитросомонас: 2NH3 + 302 = 2HN02 + 2H20. Аммиак — результат гниения белков в почве. Во-вторых, ведется окисление азотистой кислоты до азотной бактериями нитробактер: 2HN02 + 02 = 2HN03.
6. Водородные бактерии окисляют водород, который безостановочно образуется при анаэробном (без присутствия кислорода) разложении микроорганизмами почвы ряда органических остатков по формуле 2Н2 + 02 = 2Н20.
7. Хемосинтезирующие бактерии ведут окисление соединений железа и марганца. Благодаря их активной работе на дне морей и болот накапливаются огромные запасы отложенных марганцевых и железных руд. О деятельности данных бактерий по накоплению руд говорил основатель биогеохимии В. И. Вернадский.
egevideo.ru
Темновая и световая фаза фотосинтеза. Где протекает световая фаза фотосинтеза? :: SYL.ru
Фотосинтез представляет собой процесс переработки воды и углекислого газа с использованием световой энергии или без нее. Он характерен для растений. Рассмотрим далее, что собой представляют темновая и световая фаза фотосинтеза.

Общие сведения
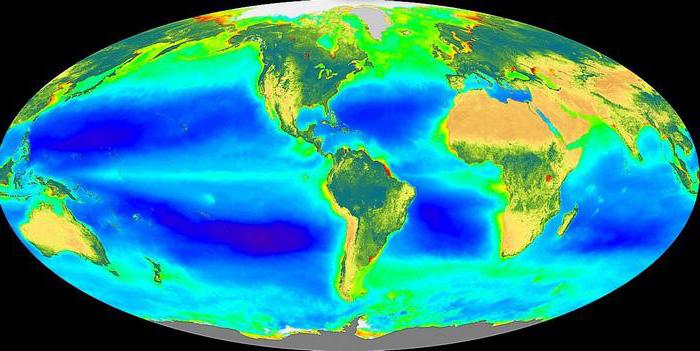
Органом фотосинтеза у высших растений является лист. В качестве органоидов выступают хлоропласты. В мембранах их тилакоидов присутствуют фотосинтетические пигменты. Ими являются каротиноиды и хлорофиллы. Последние существуют в нескольких видах (а, с, b, d). Главным из них считается а-хлорофилл. В его молекуле выделяется порфириновая «головка» с атомом магния, расположенным в центре, а также фитольный «хвост». Первый элемент представлен в виде плоской структуры. «Головка» является гидрофильной, поэтому располагается на той части мембраны, которая направлена к водной среде. Фитольный «хвост» является гидрофобным. За счет этого он удерживает хлорофилльную молекулу в мембране. Хлорофиллами поглощается сине-фиолетовый и красный свет. Они также отражают зеленый, за счет чего растения имеют характерный для них цвет. В мембранах тилактоидов молекулы хлорофилла организованы в фотосистемы. Для синезеленых водорослей и растений характерны системы 1 и 2. Фотосинтезирующие бактерии имеют только первую. Вторая система может разлагать Н2О, выделять кислород.
Световая фаза фотосинтеза
Процессы, происходящие в растениях, отличаются сложностью и многоступенчатостью. В частности, выделяют две группы реакций. Ими являются темновая и световая фаза фотосинтеза. Последняя протекает при участии фермента АТФ, белков, переносящих электроны, и хлорофилла. Световая фаза фотосинтеза происходит в мембранах тилактоидов. Хлорофилльные электроны возбуждаются и покидают молекулу. После этого они попадают на внешнюю поверхность мембраны тилактоида. Она, в свою очередь, заряжается отрицательно. После окисления начинается восстановление молекул хлорофилла. Они отбирают электроны у воды, которая присутствует во внутрилакоидном пространстве. Таким образом, световая фаза фотосинтеза протекает в мембране при распаде (фотолизе): Н2О + Q света → Н+ + ОН—
Ионы гидроксила превращаются в реакционноспособные радикалы, отдавая свои электроны:
ОН— → •ОН + е—
•ОН-радикалы объединяются и образуют свободный кислород и воду:
4НО• → 2Н2О + О2.
При этом кислород удаляется в окружающую (внешнюю) среду, а внутри тилактоида идет накопление протонов в особом «резервуаре». В результате там, где протекает световая фаза фотосинтеза, мембрана тилактоида за счет Н+ с одной стороны получает положительный заряд. Вместе с этим за счет электронов она заряжается отрицательно.
Фосфирилирование АДФ
Там, где протекает световая фаза фотосинтеза, присутствует разность потенциалов между внутренней и наружной поверхностями мембраны. Когда она достигает 200 мВ, начинается проталкивание протонов сквозь каналы АТФ-синтетазы. Таким образом, световая фаза фотосинтеза происходит в мембране при фосфорилировании АДФ до АТФ. При этом атомарный водород направляется на восстановление особого переносчика никотинамидадениндинуклеотидфосфата НАДФ+ до НАДФ•Н2:
2Н+ + 2е— + НАДФ → НАДФ•Н2
Световая фаза фотосинтеза, таким образом, включает в себя фотолиз воды. Его, в свою очередь, сопровождают три важнейших реакции:
- Синтез АТФ.
- Образование НАДФ•Н2.
- Формирование кислорода.
Световая фаза фотосинтеза сопровождается выделением последнего в атмосферу. НАДФ•Н2 и АТФ перемещаются в строму хлоропласта. На этом световая фаза фотосинтеза завершается.
Другая группа реакций
Для темновой фазы фотосинтеза не нужна световая энергия. Она идет в строме хлоропласта. Реакции представлены в виде цепочки последовательно происходящих преобразований поступающего из воздуха углекислого газа. В итоге образуются глюкоза и прочие органические вещества. Первой реакцией является фиксация. В качестве акцептора углекислого газа выступает рибулозобифосфат (пятиуглеродный сахар) РиБФ. Катализатором в реакции является рибулозобифосфат-карбоксилаза (фермент). В результате карбоксилирования РиБФ формируется шестиуглеродное неустойчивое соединение. Оно практически мгновенно распадается на две молекулы ФГК (фосфоглицериновой кислоты). После этого идет цикл реакций, где она через несколько промежуточных продуктов трансформируется в глюкозу. В них используются энергии НАДФ•Н2 и АТФ, которые были преобразованы, когда шла световая фаза фотосинтеза. Цикл указанных реакций именуется «циклом Кальвина». Его можно представить следующим образом:
6СО2 + 24Н+ + АТФ → С6Н12О6 + 6Н2О
Помимо глюкозы, в ходе фотосинтеза образуются прочие мономеры органических (сложных) соединений. К ним, в частности, относят жирные кислоты, глицерин, аминокислоты нуклеотиды.

С3-реакции
Они представляют собой тип фотосинтеза, при котором в качестве первого продукта образуются трехуглеродные соединения. Именно он описан выше как цикл Кальвина. В качестве характерных особенностей С3-фотосинтеза выступают:
- РиБФ является акцептором для углекислого газа.
- Реакция карбоксилирования катализирует РиБФ-карбоксилаза.
- Образуется шестиуглеродное вещество, которое впоследствии распадается на 2 ФГК.
Фосфоглицериновая кислота восстанавливается до ТФ (триозофосфатов). Часть из них направляется на регенерацию рибулозобифосфата, а остальная — превращается в глюкозу.
С4-реакции
Для этого типа фотосинтеза характерно появление четырехуглеродных соединений в качестве первого продукта. В 1965 году было выявлено, что С4-вещества появляются первыми у некоторых растений. Например, это было установлено для проса, сорго, сахарного тростника, кукурузы. Эти культуры стали именовать С4-растениями. В следующем, 1966-м, Слэк и Хэтч (австралийские ученые) выявили, что у них почти полностью отсутствует фотодыхание. Также было установлено, что такие С4 растения намного эффективнее осуществляют поглощение углекислого газа. В результате путь трансформации углерода в таких культурах стали именовать путем Хэтча-Слэка.

Заключение
Значение фотосинтеза очень велико. Благодаря ему из атмосферы ежегодно поглощается углекислый газ в огромных объемах (миллиардами тонн). Вместо него выделяется не меньшее количество кислорода. Фотосинтез выступает в качестве основного источника формирования органических соединений. Кислород участвует в образовании озонового слоя, обеспечивающего защиту живых организмов от воздействия коротковолновой УФ-радиации. В процессе фотосинтеза лист поглощает только 1% всей энергии света, падающего на него. Его продуктивность находится в пределах 1 г органического соединения на 1 кв. м поверхности за час.
www.syl.ru
Темновая фаза фотосинтеза
Кислород – важнейшая составляющая существования всего живого на Земле. Удивительно, но этот элемент на нашей планете, хоть его концентрация в воздухе по данным некоторых ученых неумолимо уменьшается, является восполнимым запасом. Еще более поразительным кажется тот факт, что синтезируется он из более чем доступных ресурсов – воды, солнечного света и углекислого газа. И осуществляют этот чудесный процесс растения.
Конечно, речь идет о фотосинтезе – удивительном творении природы. Несмотря на то, что ученые досконально изучили этот вопрос, повторить этапы фотосинтеза в лабораторных условиях нереально по сей день.
Этот процесс принято делить на два этапа:
- Световая фаза фотосинтеза.
- Темновая фаза фотосинтеза.
Из их названия вполне ясно, что первая часть процесса протекает на свету, то есть при участии солнечных лучей. Происходит она только в зеленых листьях растений, поскольку те содержат хлоропласты – особые элементы, в мембранах которых осуществляется синтез АТФ – молекулы, в которой запасается энергия.
При попадании фотонов солнечного света на листья растений, содержащих хлорофилл, происходит превращение энергии солнечного света в энергетические молекулы АТФ, уже упомянутые выше. Кроме того, благодаря отщеплению двух атомов водорода от молекулы воды (что также происходит при помощи солнечного света) образуется молекула НАДФ. Разложенная молекула воды, лишенная двух атомов водорода, остается со свободным кислородом, который и поступает в атмосферу. Таким образом, продукты фотосинтеза в световой фазе – это:
- кислород;
- энергетическая молекула АТФ;
- атомарный водород НАДФ Н2.
Любопытно, что образование кислорода в этом процессе вовсе не является конечной целью. Скорее, это побочный эффект. Далее происходит темновая фаза фотосинтеза, или хемосинтез, в котором принимают непосредственное участие продукты первой фазы. Рассмотрим его поподробнее.
Действительно, целью процесса не является образование кислорода. Темновая фаза фотосинтеза протекает в иной части листа – в стромах его хлоропластов. По окончанию световой фазы растение успевает запастись внушительным количеством энергетических молекул – АТФ и НАДФ Н2, следовательно, участие света больше не является необходимым. Именно с помощью этих молекул происходит синтез органических элементов. Логично, что задача энергетической молекулы АТФ – поставка энергии для осуществления процессов синтеза, в то время как роль НАДФ Н2 – восстановление.
В начале этой фазы молекула восстановителя окисляется, благодаря чему исчезают два атома водорода, что на выходе дает чистую молекулу НАДФ. В то же время АТФ отдает остаток фосфорной кислоты, превращаясь в АДФ. Эти два процесса происходят в матриксе листа. Вновь полученные молекулы после этого возвращаются в грани листьев, что дает возможность повторить весь процесс световой фазы. Однако и это не является ключевым процессом фотосинтеза, мы лишь обозначили цикличность и последовательность операций, происходящих в листьях.
Конечным продуктом данной фазы становится глюкоза – органическое соединение, относимое к простым сахарам. Впервые подробно описать синтез этой молекулы смог Мелвин Кальвин. Выяснилось, что обе молекулы, рассмотренные в рамках световой фазы, – энергетическая и восстановитель – участвуют в процессах синтеза. Кроме того, важными элементами для образования простых сахаров являются 6 молекул углекислого газа (CO2), 24 атомов водорода, 6 молекул воды:
6СО2 + 24Н + АТФ С6Н12О6 + 6Н2O.
Темновая фаза фотосинтеза важна растениям потому, что кроме глюкозы в этот период образуются различные аминокислоты, нуклеотиды, жирные кислоты и глицерин.
Фотосинтез – в высшей степени уникальный природный процесс. Он не только является залогом поддержания постоянного уровня кислорода в атмосфере и озоновом слое, но и являет собой совершенство природы, когда из неорганических элементов создаются органические.
fb.ru