Как правильно писать русские имена английскими буквами?
- Переводчики художественной литературы обычно стараются перевести «говорящие», «цветочные» и «назидательные» имена. Девушка по имени Роза станет Rose, Веру назовут Faith. Такой перевод допустим для более глубокого понимания замысла автора произведения. В обычной жизни было бы странно назвать Надежду словом Hope.
- Чтобы погрузить учеников в языковую среду, например в школе или на языковых курсах, им подбирают похожие английские имена. Елена будет Helen, Катя — Kate, Михаил — Michael, Маша — Mary.
- Для официально-делового стиля применяется метод транслитерации. О нем поговорим ниже.
Оставить заявку на обучение, вы можете здесь
Транслитерация
Транслитерация — передача знаков одного языка символами другого.
Необходимо запомнить, что в деловой переписке и в официальных документах имена собственные — имя, отчество, фамилия — не переводятся, к ним не подбираются англоязычные аналоги. Требуется записать слово латиницей. Но сложность заключается в том, что нет одинаковых правил, четких норм транслитерации. Написание имени Евгения можно увидеть как Evgenia, Yevgenia, Yevgeniya. Однако общие требования к транслитерации все же возможно определить.
Общие правила написания
Транслитерация с кириллицы на латиницу выглядит следующим образом:
A – A | И – I | С – S | Ъ – не обозначаем |
Б – B | Й – Y | Т – T | Ы – Y |
В – V | К – K | У – U | Ь – не обозначаем |
Г – G | Л – L | Ф – F | Э – E |
Д – D | М – M | Х – KH | Ю – YU |
Е – E (YE) | Н – N | Ц – TS | Я – YA |
Ё – E(YE) | О – O | Ч – CH | |
Ж – ZH | П – P | Ш – SH | |
З – Z | Р – R | Щ – SHCH |
Частные правила транслитерации
- Когда нам необходимо написать русские окончания в именах собственных, тогда правила такие:
- -ай — -ai
- -ая — -aya
- -ий — -y(iy)
- -ей — -ei
- -ия — -ia
- -ой — -oy(i)
- -ый — -yi
- Сочетание двух согласных букв «дж» пишем так: j, dj или dzh.

- «Кс» лучше передать сочетанием ks вместо x, чтобы слово звучало более правильно.
- Буквы Е и Э записываем как Е, но если она находится в начале слова, или после Ъ, Ь знаков, или после гласной, то ее пишем как YE
- Буква Ё, как правило, пишется как Е, но если мы хотим выделить его именно как звук Ё, тогда применяем сочетание YO — Pyotr, Fyodor.

Особенности транслитерации в паспорте
Правила и нормы транслитерации для загранпаспортов видоизменялись достаточно часто. На сегодняшний день все государственные организации подчиняются приказу №889 МВД России, согласно которому транслитерация имен и фамилий для загранпаспорта будет следующей:
Если вы откроете свой загранпаспорт, то увидите, что в нем не написано отчество. Дело в том, что такое понятие, как «отчество» (patronymic name), отсутствует в английской культуре. Однако многие жители европейских стран, а также жители англоязычных стран имеют
Однако многие жители европейских стран, а также жители англоязычных стран имеют
Достаточно часто может возникнуть необходимость проговорить свое ФИО по-английски. В таких ситуациях мы говорим имя, затем отчество, а потом — фамилию. Например: Ivan Sergeyevich Turgenev.
Когда мы заполняем документ, в графе First name указываем имя, а в графе Last name (Second name) пишем фамилию. Отчество записывать не надо.
Иногда вместо First name можно увидеть Given name (forename) или Christian name — это все синонимы, обозначающие имя, данное при рождении.
Если вы сомневаетесь, как написать свое имя на английском, то ниже в таблице мы привели примеры, как пишутся русские имена на английском и их уменьшительные формы.
Мужские имена
Русское имя | Транслитерация | Сокращенное имя | Транслитерация |
Александр | Aleksandr, Alexander | Саша | Sasha |
Алексей | Aleksey, Alexey | Алёша | Alyosha |
Альберт | Albert | Алик | Alik |
Анатолий | Anatoly, Anatoliy | Толя | Tolya |
Андрей | Andrey, Andrei | Андрюша | Andryusha |
Антон | Anton | Антоша | Antosha |
Аркадий | Arkady, Arkadiy | Аркаша | Arkasha |
Артём | Artem, Artyom | Тёма | Tyoma |
Артур | Artur | ||
Архип | Arkhip | ||
Афанасий | Afanasii, Afanasiy | Афоня | Afonia |
Борис | Boris | Боря | Borya |
Бронислав | Bronislav | ||
Вадим | Vadim | Вадик | |
Валентин | Valentin | Валя | Valya |
Валерий | Valeriy | Валера | Valera |
Василий | Vasily, Vasiliy | Вася | Vasya |
Виктор | Viktor, Victor | Витя | Vitya |
Виталий | Vitaly, Vitaliy | Виталик | Vitalik |
Владимир | Vladimir | Вова, Володя | Vova, Volodya |
Владислав | Vladislav | Влад | Vlad |
Всеволод | Vsevolod | Сева | Seva |
Вячеслав | Vyacheslav, Viacheslav | Слава | Slava |
Геннадий | Gennady, Gennadiy | Гена | Gena |
Георгий | Georgy, Georgiy | Гоша | Gosha |
Глеб | Gleb | ||
Григорий | Grigory, Grigoriy | Гриша | Grisha |
Даниил, Данила | Daniil, Danila | Даня | Dania |
Денис | Denis | ||
Дмитрий | Dmitry, Dmitriy | Дима | Dima |
Евгений | Yevgeny, Yevgeniy | Женя | Zhenya |
Егор | Yegor, Egor | Гоша | Gosha |
Ефим | Efim | Фима | Fima |
Захар | Zakhar, Zahar | ||
Иван | Ivan | Ваня | Vanya |
Игнат | Ignat | ||
Игорь | Igor | ||
Илья | Ilya, Ilia | Илюша | Ilyusha |
Иннокентий | Innokenty, Innokentiy | Кеша | Kesha |
Кирилл | Kirill | ||
Константин | Konstantin | Костя | Kostya |
Лев | Lev | Лёва | Lyova |
Леонид | Leonid | Лёня | Lyonya |
Макар | Мakar | ||
Максим | Maksim, Maxim | Макс | Max |
Матвей | Matvey, Matvei | ||
Мирон | Miron | ||
Михаил | Mikhail | Миша | Misha |
Никита | Nikita | ||
Николай | Nikolay, Nikolai | Коля | Kolya |
Олег | Oleg | ||
Осип | Osip | ||
Павел | Pavel | Паша | Pasha |
Петр | Pyotr, Petr | Петя | Petya |
Прохор | Prokhor | Проша | Prosha |
Родион | Rodion | Родя | Rodya |
Роман | Roman | Рома | Roma |
Ростислав | Rostislav | Слава | Slava |
Святослав | Svyatoslav, Sviatoslav | Слава | Slava |
Семён | Semyon | Сеня, Сёма | Senya, Syoma |
Сергей | Sergey, Sergei | Серёжа | Seryozha |
Станислав | Stanislav | Стас | Stas |
Степан | Stepan | Стёпа | Styopa |
Тимофей | Timofey, Timofei | Тима | Tima |
Тимур | Timur, Timour | Тёма | Tyoma |
Тихон | Tikhon | Тиша | Tisha |
Фёдор | Fedor, Fyodor | Федя | Fedya |
Филипп | Filipp, Philipp | ||
Эдуард | Eduard, Edward | Эдик | Edik |
Юлиан | Yulian | ||
Юрий | Yuri, Yuriy, Yury | Юра | Yura |
Яков | Yakov, Iakov | Яша | Yasha |
Ян | Yan, Ian | ||
Ярослав | Yaroslav | Ярик, Слава | Yarik, Slava |
Женские имена
Русское имя | Транслитерация | Сокращенное имя | Транслитерация |
Алевтина | Alevtina | Аля | Alia |
Александра | Aleksandra, Alexandra | Саша | Sasha |
Алёна | Alyona | ||
Алиса | Alisa | ||
Алина | Alina | Аля | Alya |
Алла | Alla | ||
Альбина | Albina | ||
Анастасия | Anastasia, Anastasiya | Настя | Nastya |
Ангелина | Angelina | Лина | Lina |
Анжела | Anzhela, Angela | ||
Анна | Anna | Аня | Anya |
Антонина | Antonina | Тоня | Tonya |
Анфиса | Anfisa | ||
Валентина | Valentina | Валя | Valya |
Валерия | Valeria, Valeriya | Лера | Lera |
Варвара | Varvara | Варя | Varya |
Василина | Vasilina | Вася | Vasia |
Вера | Vera | ||
Вероника | Veronika, Veronica | Ника | Nika |
Виктория | Viktoria, Viktoriya, Victoria | Вика | Vika |
Владислава | Vladislava | Влада | Vlada |
Галина | Galina | Галя | Galya |
Дарина | Darina | Даша | Dasha |
Дарья | Darya | Даша | Dasha |
Диана | Diana | ||
Дина | Dina | ||
Ева | Eva | ||
Евгения | Yevgenia, Yevgeniya, Evgenia | Женя | Zhenya |
Екатерина | Yekaterina, Ekaterina | Катя | Katya |
Елена | Yelena, Elena | Лена | Lena |
Елизавета | Yelizaveta, Elizaveta | Лиза | Liza |
Жанна | Zhanna | ||
Зинаида | Zinaida | Зина | Zina |
Зоя | Zoya | ||
Инга | Inga | ||
Инна | Inna | ||
Инесса | Inessa | ||
Ирина | Irina | Ира | Ira |
Кира | Kira | ||
Клавдия | Klavdiia, Klavdiya | Клава | Klava |
Клара | Klara, Clara | ||
Кристина | Kristina | ||
Ксения | Ksenia, Kseniya | Ксюша | Ksiusha |
Лариса | Larisa | Лара | Lara |
Лидия | Lidia, Lidiya | Лида | Lida |
Лилия | Lilia, Liliya | Лиля | Lilya |
Любовь | Lyubov, Liubov | Люба | Lyuba |
Людмила | Lyudmila, Liudmila | Люда | Lyuda |
Майя | Maya, Maia | ||
Маргарита | Margarita | Рита | Rita |
Мария | Maria, Mariya | Маша | Masha |
Марина | Marina | ||
Марта | Marta | ||
Надежда | Nadezhda | Надя | Nadya |
Наталья, Наталия | Natalya, Natalia, Nataliya | Наташа | Natasha |
Нина | Nina | ||
Оксана | Oksana, Oxana | Ксюша | Ksyusha |
Олеся | Olesya, Olesia | Леся | Lesya |
Ольга | Olga | Оля | Olya |
Полина | Polina | Поля | Polya |
Раиса | Raisa | Рая | Raya |
Регина | Regina | ||
Рената | Renata | ||
Римма | Rimma | ||
Роза | Rosa | ||
Светлана | Svetlana | Света | Sveta |
Снежана | Snezhana | ||
София, Софья | Sofya, Sofia | Соня | Sonya |
Таисия | Taisiya | Тая | Taya |
Тамара | Tamara | Тома | Toma |
Татьяна | Tatyana, Tatiana | Таня | Tanya |
Ульяна | Ulyana, Uliana | Уля | Ulya |
Фаина | Faina | Фая | Faya |
Эвелина | Evelina | ||
Элла | Ella | ||
Юлиана | Yuliana | Юля | Yulya |
Юлия | Yulia, Yuliya | Юля | Yulya |
Яна | Yana | ||
Ярослава | Yaroslava | Слава | Slava |
Русские имена представляют ту еще задачку, когда нужно заполнить документ на английском и написать свое имя латиницей. Мы постарались дать вам всю самую важную информацию, чтобы в следующий раз, когда вы закажете очередную классную посылку с американского Ebay, заказ пришел именно к вам.
Мы постарались дать вам всю самую важную информацию, чтобы в следующий раз, когда вы закажете очередную классную посылку с американского Ebay, заказ пришел именно к вам.
Транскрипция… учить или не учить? Не учить!
Транскрипция… учить или не учить? Не учить!15.10.2020
Даже в наше время при обучении английскому языку встает вопрос о необходимости изучения
транскрипции.Почему многим родителям кажется правильным учить транскрипцию? Они думают, что эти знания научат детей читать. Но это не так.
- Ведь многие родители сами когда-то учили транскрипцию в школе и считают эти знания обязательными для своих детей. Однако прогресс не стоит на месте и вносит свои коррективы в образовательный процесс.
- В советское время транскрипция помогала узнать, как произносится незнакомое слово. И тогда это был чуть ли не единственный способ. Транскрипция была нужна не для чтения, а лишь для пользования словарем.
- В наше же время в распоряжении у учеников и студентов великое множество различных ресурсов, благодаря которым возможно узнать правильное произношение той или иной лексики. Отдельного внимание заслуживает изучение транскрипции в младшем школьном или дошкольном возрасте. Стоит понимать, что маленького ребенка транскрипция однозначно только запутает. Дошкольник или первоклассник, который только недавно выучил русский алфавит, изучает английский алфавит — это уже две знаковые системы, которые он должен держать в голове. И тут вводится третья знаковая система, которая нужна, чтобы расшифровать вторую. Представляете, что творится в голове у ребенка?
И это еще не все!
Наиболее популярная и эффективная методика в наши дни – коммуникативная.
- Транскрипционный алфавит почти в два раза длиннее английского алфавита. Часто дети начинают путать буквы и звуки, потому что запоминают именно транскрипционный вариант написания слова. От этого следуют и дальнейшие проблемы с чтением.
Она позволяет отказаться от изучения транскрипции.Можно использовать онлайн-словари, произносящие слово голосом носителя языка, для того, чтобы запомнить произношение. Достаточно просто повторить слово за диктором словаря. Если у ребенка не очень хороший музыкальный слух, ему может быть сложно распознать правильно все звуки на слух в онлайн-словаре, в таком случае лучше практиковаться с преподавателем, который поправит и создаст ситуацию, когда слово повторяется много раз.
Еще один способ научить ребенка читать без транскрипции – метод целых слов.Механизм такой — ребенок не разбирает слово на звуки, он соотносит зрительный образ целого слова с его звучанием (благодаря этому, метод целых слов можно начинать использовать параллельно с изучением алфавита и звуков, не дожидаясь, пока все буквы будут изучены). Потом, видя изученное слово в тексте, ребенок вспоминает, как оно звучит и может это слово прочитать. Способ хорош тем, что в английском чуть ли не половина слов — исключения, которые читаются не по правилам и иного способа выучить эти слова, кроме как запомнить просто нет.
В процессе обучения главное не отбить у ребенка желание учиться и сделать условия учебы максимально комфортными и интересными.В Языковой школе ILS мы слышим наших учеников и используем самые эффективные методы обучения.
ХОТИТЕ, ЧТОБЫ ВАШ РЕБЕНОК УЧИЛСЯ С УДОВОЛЬСТВИЕМ И ПРИНОСИЛ ДОМОЙ ПЯТЕРКИ?



ОБРАЩАЙТЕСЬ К МЕНЕДЖЕРАМ ЯЗЫКОВОЙ ШКОЛЫ ILS ?+7 495 215 58 28
Поделиться статьей
404 — Документ не найден
Филологическая олимпиада: старт заочного этапа (02.12.21)
1 декабря начинается заочный этап Филологической олимпиады для школьников 5-11 классов «Юный словесник». Тематическое направление 2021-2022 учебного года: «Тема дружбы в русском языке, литературе и культуре». Ответы принимаются с 15 декабря 2021 года по 15 января 2022 года …
Новости
Внимание! Интересная и перспективная вакансия на телевидении! (26. 01.22)
01.22)
ГТРК «Иртыш» срочно ищет продюссера новостей! …
Объявления
О функционировании факультета в период с 9 по 27 февраля 2022 года (07.02.22)
учебный процесс с 09.02.2022 по 27.02.2022 г. осуществляется с применением электронного обучения и дистанционных образовательных технологий для направлений Филология, Туризм, БИД, РиСО, РКИ, Новейшая филология и в смешанном режиме в соответствии с расписанием занятий для направления Журналистика. …
Конференция «Актуальные проблемы русского языка и методики его преподавания: традиции и инновации» (07.02.22)
15 апреля 2022 г. состоится XIX Всероссийская научно-практическая конференция молодых ученых «Актуальные проблемы русского языка и методики его преподавания: традиции и инновации» …
Требуются учителя русского языка и литературы (27.01.22)
БОУ СОШ № 51 г. Омска требуются учителя русского языка и литературы (2 ставки). Возможно трудоустройство студентов. …
Возможно трудоустройство студентов. …
Конференция «Актуальные проблемы медиапространства: от теоретических аспектов до практической реализации» (14.01.22)
Российский университет дружбы народов приглашает студентов и преподавателей принять участие в международной научно-практической конференции «Актуальные проблемы медиапространства: от теоретических аспектов до практической реализации» …
Транслитерация (русские имена и фамилии по-английски)
Правила транслитерации. Как русские имена и фамилии пишутся по-английски.
Когда нужно записать имя (или фамилию) на другом языке, возможно применение транскрипции, когда слово переписывают буквами другого языка, стараясь передать произношение слова на языке-источнике, но обычно используется транслитерация, когда буквы одного языка заменяют буквами другого языка по определенному правилу.
*** Для тех, кто хотел знать, как слово «Яндекс» пишется по-английски: yandex (сайт http://yandex. ru)
ru)
ru — русская буква, en — по-английски
| ru | en | Пример транслитерации | Соответствующее английское имя | |
| а | a | Anna, Anya Anastasia Aleksandr Aleksei, Aleksey Andrey Artur | Анна, Аня Анастасия Александр Алексей Андрей Артур | Ann [æn] — Alexander[ˌælɪɡˈzɑːndə] — Andrew [ˈændruː] Arthur [ˈɑːθə] |
| б | b | Boris, Borya | Борис, Боря | |
| в | v | Varvara Vladimir, Vova Valery Vitya | Варвара Владимир, Вова Валерий Витя | Barbara[ˈbɑːb(ə)rə] — — — |
| г | g | Grigory, Grisha | Григорий, Гриша | |
| д | d | Dmitry, Dima | Дмитрий, Дима | |
| е | e, ye | Elena Egor Evgeniy Yeltsin Boris Nikolaevich | Елена Егор Евгений Ельцин Борис Николаевич | Helen[ˈhɛlən] — Eugene [ˈjuːdʒiːn] — — |
| ё | yo | Yolkin | Ёлкин | |
| ж | zh | Zhenya | Женя | |
| з | z | Zinaida, Zina | Зинаида, Зина | |
| и | i | Irina, Ira Igor | Ирина, Ира Игорь | Irene [ˈaɪriːn] — |
| й | y | Valery Leontyev | Валерий Леонтьев | |
| к | c, k | Cirill Kolya Victoria, Vica | Кирилл Коля Виктория, Вика | |
| л | l | Lidia, Lida, Lena | Лидия, Лида, Лена | |
| м | m | Maria, Masha Mikhail, Misha Margarita | Мария, Маша Михаил, Миша Маргарита | Mary [ˈmɛərɪ] Michael [ˈmaɪk(ə)l] Margaret [ˈmɑːɡ(ə)rət] |
| н | n | Nicolay Natasha Nina | Николай Наташа Нина | Nicholas [ˈnɪk(ə)ləs] — — |
| о | o | Olga, Olya Oleg | Ольга, Оля Олег | |
| п | p | Piotr, Petya Pavel Polyna, Polina | Пётр, Петя Павел Полина | Peter [ˈpiːtə] Paul [pɔːl] Paulina [pɔːˈliːnə] |
| р | r | Rita Roman | Рита Роман | |
| с | s | Sergey, Seryozha Sveta Sasha | Сергей, Сережа Света Саша | |
| т | t | Tatyana, Tanya Tamara | Татьяна, Таня Тамара | |
| у | u | Ulyana | Ульяна | |
| ф | f | Fyodor | Федор | Theodore [ˈθiːədɔː] |
| х | kh | Svetlana Khorkina | Светлана Хоркина | |
| ц | ts | Tseitlin | Цейтлин | |
| ч | ch | Chulpan Hamatova | Чулпан Хаматова | |
| ш | sh | Shura Vasily Shukshyn | Шура Василий Шукшин | |
| щ | shch | Shchukin Boris Vasilyevich Tatishchev Vasily Nikitich | Щукин Борис Васильевич Татищев Василий Никитич | |
| ъ | “ | |||
| ы | y | Krylov Ivan Andreevich | Крылов Иван Андреевич | |
| ь | ‘ | |||
| э | e | Elen Ella Andrey Eshpay | Элен Элла Андрей Эшпай | Helen[ˈhɛlən] — |
| ю | yu | Yulia, Yulya Yuriy, Yura | Юлия, Юля Юрий, Юра | Julia [ˈdʒuːlɪə] — |
| я | ya | Yakov | Яков | Jacob [ˈdʒeɪkəb] |
На практике многие имена пишут по-разному, например,
Василий — Vasily Vasiliy, Vasili, Vasilii
Юрий — Yury Yuri Yuriy
Вот типичная цитата: «. ..у меня в загранпаспорте написали Dmitry. Я Юрьевич, так в загранпаспорте у отца вообще Youry…»
..у меня в загранпаспорте написали Dmitry. Я Юрьевич, так в загранпаспорте у отца вообще Youry…»
(еще и по правилам французского языка!)
А вот правила для транслитерации, используемые Сбербанком для написания имен и фамилий на пластиковых картах:
| А -> A | Б -> B | В -> V |
| Г -> G | Д -> D | Е -> E |
| Ё -> E | Ж -> ZH | З -> Z |
| И -> I | Й -> Y | К -> K |
| Л -> L | М -> M | Н -> N |
| О -> O | П -> P | Р -> R |
| С -> S | Т -> T | У -> U |
| Ф -> F | Х -> KH | Ц -> TS |
| Ч -> CH | Ш -> SH | Щ -> SHCH |
| Ъ -> | Ы -> Y | Ь -> |
| Э -> E | Ю -> YU | Я -> YA |
* русские гласные е,ё, ю, я пишутся :
через y: ye, yo, yu, ya — после гласной и в начале слова;
через i: ie, io, iu, ia — после согласной
Например,
АЛЕКСАНДР ~ ALEXANDER
ВЯЧЕСЛАВ ~ VJATCHESLAV
ВИКТОР ~ VICTOR
КУЗЬМА ~ KOUZMA
ЛЮБОВЬ ~ LIUBOV
ЛЮДМИЛА ~ LIUDMILA
НАДЕЖДА ~ NADEZDA
ФИЛИПП ~ PHILIPP
ЮЛИЯ ~ JULIA
ЮРИЙ ~ YURI
ЯКОВ ~ IAKOV
Наверное, самым авторитетным должен быть сайт Президента России. На сайте
На сайте
http://eng.kremlin.ru/ имя Дмитрия Медведева по-английски пишется Dmitry Medvedev.
Полезные ссылки:
Русские имена: значение и происхождение: kurufin.narod.ru/html/rus.html
Перевод русских букв в английские (онлайн): fotosav.ru/services/transliteration.aspx
Дополнительно: в разделе «Справочник» также:
Зульфикар Али Бхутто осуждает Совет Безопасности ООН
ОРГАНИЗАЦИЯ ОБЪЕДИНЕННЫХ НАЦИЙ, Нью-Йорк, 15 декабря — Министр иностранных дел Пакистана Зульфикар Алл Бхутто со слезами на лице вышел сегодня из Совета Безопасности, обвинив его в «узаконивании агрессии».
Выйдя из комнаты, он сказал: «Я ненавижу это тело. Я не хочу снова видеть их лица. Я лучше вернусь в разрушенный Пакистан».
Затем в сопровождении семи членов его делегации с мрачными лицами, включая постоянного представителя Ага Шаха!Бхутто прошел по устланному ковром главному залу мимо суетящихся группок удивленных дипломатов и был унесен туманным дождем.
Г-н Бхутто не указал непосредственную причину своих действий. Но, говоря о «тактике проволочек» и «обструкции», он, по-видимому, имел в виду последовательное наложение вето Советским Союзом — сторонником Индии — на резолюции, призывающие к выводу войск.
Прощальные слова г-на Бхутто Совету, прежде чем он разорвал свои записи, отодвинул стул и встал, были следующими:
«Г-н.Президент, я не крыса. Я никогда в жизни не крысил. Я сталкивался с покушениями, я сталкивался с тюремным заключением. Сегодня я не сдаюсь, а покидаю ваш Совет Безопасности.
«Я считаю постыдным для себя и своей страны оставаться здесь дольше. Навязывать любое решение, иметь договор хуже Версальского, легализовать агрессию, легализовать оккупацию — я не буду его участником. Мы будем бороться. Моя страна слушает меня.
«Зачем мне тратить время здесь, в Совете Безопасности? Я не буду участником позорной капитуляции части моей страны.Вы можете взять свой Совет Безопасности; а, вот и ты. Я ухожу».
Я ухожу».
Возобновление дебатов
Делегаты вокруг круглого стола бесстрастно смотрели на него, когда он выходил из зала. Несколько мгновений спустя президент Совета Исмаэль Б. ТейлорКамера предоставил слово Рашиду Дриссу из Туниса, и дебаты продолжились, как вчера и позавчера.
Проходя по коридору, г-н Бхутто заявил, что Пакистан не разрывает отношений с Советом или Организацией Объединенных Наций.
«Посол Шахи будет доступен», — сказал он.
Совет собрался дважды
Вечером Совет снова собрался дважды. Китайские и советские делегаты, выступавшие главными докладчиками на первом вечернем заседании, обменялись резкими обвинениями в политике большой державы и идеологическом предательстве.
Но во втором сеансе, который начался незадолго до 11 вечера, появился намек на то, что тупик большой мощности завтра может выйти из тупика.
Делегаты из Великобритании и Франции, сэр Колин Кроу и Жак Костюшко-Моризе, представили совместное предложение, основные положения которого призывали к прекращению огня и всеобъемлющему политическому урегулированию между Пакистаном и сепаратистскими повстанцами.
Яков А. Малик из Советского Союза также представил резолюцию, призывающую к прекращению огня и «одновременному» политическому урегулированию. Западные дипломаты связали его краткое и невнятное заявление с тем фактом, что сводки новостей, поступившие в зал Совета вечером, предвещали неминуемое падение Дакки.
Многие из этих дипломатов предполагали, что Советский Союз, который поддерживал Индию, позволит Совету действовать после падения Дакки.
Около полуночи Совет прервался без голосования до завтра.
Первоначально Совет был созван в спешке в 12:30. по просьбе мистера Бхутто.
Сначала казалось, что г-н Бхутто ушел в гневе по поводу британо-французской инициативы. Во время заседания Совета он косвенно обвинил в трусости две делегации, которые ранее воздерживались от всех резолюций, призывающих к прекращению огня и выводу войск.
— Галльская логика и британский опыт, чем бы они ни были, — усмехнулся он. «Помните, что нейтрального животного не бывает. Я уважаю россиян за четкую позицию». Но после своего ухода г-н Бхутто заявил, что его делегация не отвергла британо-французский проект.
Я уважаю россиян за четкую позицию». Но после своего ухода г-н Бхутто заявил, что его делегация не отвергла британо-французский проект.
Г-н Бхутто, у которого часто срывался голос, сообщил Совету, что его 11-летний сын позвонил ему из Пакистана и сказал: «Не возвращайтесь с документом о капитуляции».
«Прийти сюда необходимо»
«Я чувствовал, что мне необходимо приехать сюда и добиться справедливости от Совета Безопасности», — сказал г-н Бхутто.«Но я должен сказать, что Совет Безопасности отказал моей стране в этом правосудии. С того момента, как я прибыл, мы были пойманы тактикой проволочек.
«Боюсь, Совет Безопасности преуспел в искусстве пиратства. С некоторым цинизмом я наблюдал, как вчера целый час был потрачен впустую на то, будут ли участники готовы встретиться. в 9:30 утра или кровать и завтрак требовали, чтобы они встретились в 11 часов утра.
«Представитель Сомали называл население Восточного Пакистана 56 миллионами, но позже поправился, назвав 76 миллионов. Если бы он подождал еще несколько дней, ему не нужно было бы исправляться, потому что умирают миллионы.
Если бы он подождал еще несколько дней, ему не нужно было бы исправляться, потому что умирают миллионы.
«Памятник вето»
«Нас разочаровало вето. Давайте построим памятник вето. Давайте построим памятник бессилию и недееспособности.
«Но помните библейское изречение: «Что посеешь, то и пожнешь». Сегодня мы охотничьи свинки, но найдутся и другие».
Обращаясь к Якову А. Малику, советскому делегату, г-н Бхутто сказал: «Выбрасываете грудь и бьете по столу.Вы говорите не как товарищ Малик, а как царь Малик. Я рад, что ты улыбаешься. Нет, мое сердце разрывается».
Позже в своем номере в отеле «Пьер» г-н Бхутто сказал, что он выступает за переговоры с сепаратистами Восточного Пакистана, но при одном условии, а именно, чтобы Пакистан был признан как субъект. Он указал, что связь между Ист-Эндом и Западным Пакистаном может быть очень слабой.
Как могли вестись такие переговоры, если Дакка пала? его спросили. Он ответил: «Я думаю, что сепаратистские лидеры сочтут в своих интересах не закрывать дверь перед Пакистаном. Они захотят поговорить как с Индией, так и с Пакистаном».
Они захотят поговорить как с Индией, так и с Пакистаном».
Г-н Бхутто добавил, что он принимает британо-французскую резолюцию в качестве основы для переговоров при условии, что в ней не указывается, что одним из партнеров по переговорам является Бангладеш или Бенгальская нация, независимая страна, провозглашенная бенгальскими повстанцами на Востоке.
Примечания Советское «давление»
Г-н Бхутто сказал, что Советский Союз оказывал «огромное давление» в сообщениях не только на Пакистан, но и на другие азиатские страны с начала кризиса.Русские превзошли индийцев», — сказал он.
Он выразил опасение, что, получив полную свободу действий в Восточном Пакистане, Индия при поддержке СССР может двинуться в пакистанскую часть Кашмира, спровоцировав таким образом китайскую интервенцию.
На протяжении всего интервью прозвучало сильное предположение, что пакистанское военное правительство неспособно справиться с нынешней ситуацией и что Пакистану необходимо вернуться к народному правительству.
Во время интервью г.Бхутто позвонила его 18-летняя дочь, которая изучает государственное управление в Гарварде. Его часть разговора выглядела так:
«Я не выбежал, я вышел. Это был слишком большой фарс. Пришлось ввести немного правды. Нет. Я не вышел из себя.
Харрис Леновиц: Из книги Джейкоба Франка «Слова Господа»
У смертного одра Якоба Франка 1791Перевод польских рукописей Харриса Леновица
ПРИМЕЧАНИЕ.Восемнадцатый век был временем растущих потрясений и деконструкций и стал свидетелем перемен в сознании, которые достигли изолированных уголков Европы, далеких от твердынь Просвещения, «естественного сверхъестественного» и радикального мистицизма, которые были одними из признаков зарождающийся романтизм. Мессианское франкистское движение, затронувшее восточноевропейских евреев, повлекло за собой, как и его литературные и западные коллеги, сдвиг в языке и сопутствующих символах, которые напоминали сдвиги, возникающие также и в доминирующих культурах.
О произведении, представленном ниже, Харрис Леновиц пишет как переводчик: «Вот некоторые из высказываний Янкиева Лейвича, Якова бен Льва, называвшего себя Яковом Франком и которого некоторые называли Мудрым Яковом. Яков Франк [1726-1791] был выходцем из Подолии, Турции, Польши в ее распаде. Он путешествовал. Его отец был странствующим проповедником. Франк тоже был торговцем и говорил на всех языках: балканском, турецком, идиш, польском, ладино, с цитатами, цитатами и языковой игрой на иврите и арамейском.Он присоединился к саббатианцам, последователям мессианского движения, начатого Шабтаем Цви и Натаном из Газы [в семнадцатом веке] и продолжавшегося через Барухью Руссо [ум. 1721 г.], и временно укоротить одного мессию. Вместе с ними он обратился против Талмуда, в Зоар и вышел через Саббатейскую пору. Он добавил к этому движению кое-что: новый акцент на Богородице, переход через христианство после перехода через ислам, начало которому положил Шабтай/Натан, на пути к Исаву. Возможно, больше секса.Он стал мессией для тысяч евреев».
Возможно, больше секса.Он стал мессией для тысяч евреев».
В «словах», записанных его последователями, мини-рассказы показывают ряд преобразующих переживаний, которые пришли к нему, как стихи ватической прозы, в форме снов и видений или наблюдений, простых или нет, за людьми. события, к которым привел его образ жизни. (Дж.Р./Х.Л.)
[Некоторые из почти 3000 высказываний и видений следуют из полного перевода Леновица, ожидающего публикации, но полностью доступного на http://archive.org/stream/TheCollectionOfTheWordsOfTheLordJacobFrank/TheCollectionOfTheWordsOfTheLordJacobFrank_djvu.txt]:
1. Мне было видение в Салониках, как будто бы следующие слова были сказаны кому-то: Иди, введи Иакова Мудрого в комнаты, и когда ты с ним войдешь в первую комнату, я увещеваю тебя, чтобы все двери и ворота были открыты ему. Когда я вошел в первую комнату, мне дали розу как знак, по которому я мог пройти в следующую и так далее , последовательно, из одной комнаты в другую. И вот я летал по воздуху в сопровождении двух девушек, [подобных] чьей красоте мир никогда не видел. В этих комнатах я видел большей частью женщин и барышень. В некоторых, однако, собирались только группы учеников и учителей, и где бы мне ни говорили первое слово, я сразу схватывал из него все дело и весь смысл. Комнат этих было неисчислимое количество и в последней из них я увидел Первого [= Шабтая Цви], который тоже сидел как учитель со своими учениками, одетый в френк [= турецкая] одежда.Тот сразу спросил меня: ты ли Иаков мудрый? Я слышал, что вы сильны и храбры сердцем. Я пришел к этому пункту, но не в силах идти отсюда дальше; если хочешь, укрепись и да поможет тебе Бог, ибо очень многие предки брали на себя то бремя, шли по этой дороге, но падали. При этом он показал мне через окно этой комнаты бездну, подобную черному морю, скрытую в необыкновенной тьме, а по другую сторону этой бездны я увидел гору, высота которой, казалось, касалась облаков.На это я закричал: «Будь что будет, я пойду с божьей помощью», и так стал лететь наискось по воздуху в глубину, пока не достиг самого ее дна, где, пощупав землю, остановился.
И вот я летал по воздуху в сопровождении двух девушек, [подобных] чьей красоте мир никогда не видел. В этих комнатах я видел большей частью женщин и барышень. В некоторых, однако, собирались только группы учеников и учителей, и где бы мне ни говорили первое слово, я сразу схватывал из него все дело и весь смысл. Комнат этих было неисчислимое количество и в последней из них я увидел Первого [= Шабтая Цви], который тоже сидел как учитель со своими учениками, одетый в френк [= турецкая] одежда.Тот сразу спросил меня: ты ли Иаков мудрый? Я слышал, что вы сильны и храбры сердцем. Я пришел к этому пункту, но не в силах идти отсюда дальше; если хочешь, укрепись и да поможет тебе Бог, ибо очень многие предки брали на себя то бремя, шли по этой дороге, но падали. При этом он показал мне через окно этой комнаты бездну, подобную черному морю, скрытую в необыкновенной тьме, а по другую сторону этой бездны я увидел гору, высота которой, казалось, касалась облаков.На это я закричал: «Будь что будет, я пойду с божьей помощью», и так стал лететь наискось по воздуху в глубину, пока не достиг самого ее дна, где, пощупав землю, остановился. Идя в темноте, я наткнулся на край горы и, увидев, что из-за крутой глади горы мне трудно взобраться на нее, я был вынужден карабкаться руками и ногтями и изо всех сил, пока не достиг вершины. Как только я остановился там, до меня донесся необыкновенный запах; и там было много истинно верующих.Охваченный великой радостью, я не хотел [пока] всем телом подниматься на гору, говоря себе: я отдохну здесь немного, ибо пот лился с головы моей, как река в разливе, от мучений, которые Я выдержал, чтобы взобраться на эту гору; но когда я хорошо отдохну, тогда я поднимусь на гору навстречу всему хорошему, что там есть. Что я и сделал, я позволил своим ногам свисать и сел с телом и руками в покое на горе. Потом я поднялся на гору.
Идя в темноте, я наткнулся на край горы и, увидев, что из-за крутой глади горы мне трудно взобраться на нее, я был вынужден карабкаться руками и ногтями и изо всех сил, пока не достиг вершины. Как только я остановился там, до меня донесся необыкновенный запах; и там было много истинно верующих.Охваченный великой радостью, я не хотел [пока] всем телом подниматься на гору, говоря себе: я отдохну здесь немного, ибо пот лился с головы моей, как река в разливе, от мучений, которые Я выдержал, чтобы взобраться на эту гору; но когда я хорошо отдохну, тогда я поднимусь на гору навстречу всему хорошему, что там есть. Что я и сделал, я позволил своим ногам свисать и сел с телом и руками в покое на горе. Потом я поднялся на гору.
2.Когда я однажды заболел в Дзюрдзюве, мне приснился такой сон. Я видел необычайно красивую женщину, у которой был колодец с водой жизни и другой колодец с чистой водой, и эта [женщина] сказала мне: «Опусти ноги в воду, и ты сразу выздоровеешь». Я так и сделал и выздоровел. На том месте, где была найдена эта женщина, было широкое красивое поле, на котором она, взяв меня за руку, сказала: «Подойди, я покажу тебе мою дочь, которая еще девица», и я пошел с нею в глубину того поля, которое источало необыкновенный аромат [множества] разных цветов.Дева, красоту которой ничто в этом мире не могло описать, вышла нам навстречу там, и была она одета в польскую rubran [обтягивающую, скрученную блузу из тяжелого, обычно красного, шелка] и ее обнаженные груди были видны . Заметив это, я вдруг увидел с одного конца света на другой. Ее мать сообщила мне, что если я пожелаю взять ее в жены, то она это позволит, но я ответил, что у меня есть жена и дети.
На том месте, где была найдена эта женщина, было широкое красивое поле, на котором она, взяв меня за руку, сказала: «Подойди, я покажу тебе мою дочь, которая еще девица», и я пошел с нею в глубину того поля, которое источало необыкновенный аромат [множества] разных цветов.Дева, красоту которой ничто в этом мире не могло описать, вышла нам навстречу там, и была она одета в польскую rubran [обтягивающую, скрученную блузу из тяжелого, обычно красного, шелка] и ее обнаженные груди были видны . Заметив это, я вдруг увидел с одного конца света на другой. Ее мать сообщила мне, что если я пожелаю взять ее в жены, то она это позволит, но я ответил, что у меня есть жена и дети.
44. Моя бабушка, мать моей матери, была очень ученым астрологом.Когда я
Родился, все ведьмы собрались вокруг нашего дома и окружили его, даже их королева была там во главе. У нас в доме жила собака — помесь волка и [собачьей] суки. Этот совсем не спал, а все время лаял, ибо если бы он заснул хотя бы на мгновение, то проследили бы, чтобы он никогда не проснулся, но он бодрствовал зорко. Потом на 8-й день на обрезании окружили наш дом по-прежнему и хотели сделать что-то злое, но не могли, потому что та собака снова караула, а старая бабка своим ремеслом боролась и со злом, говоря: смотри на него. бережно, правильно воспитай его, ибо через него в мир придет новое.
Потом на 8-й день на обрезании окружили наш дом по-прежнему и хотели сделать что-то злое, но не могли, потому что та собака снова караула, а старая бабка своим ремеслом боролась и со злом, говоря: смотри на него. бережно, правильно воспитай его, ибо через него в мир придет новое.
451. 21 октября 1784 года Господь видел сон, У меня на руке было золотое кольцо, и я уронил это кольцо на зеркало, которое разбилось на мелкие осколки, Перевернув это зеркало на другую сторону, я нашел и там блестящее стекло, и точно так же браслет выпал у меня из рук и сломал другую сторону. Он сам дал толкование тому, что помощь Моя спешит прийти.
504. Во сне я видел Иисуса, окруженного священниками, сидящего у источника живой и чистой воды.Я заметил, что эта весна ушла от них и пришла ко мне.
748. Я видел сон, как будто я был в церкви, совершенно голый, если не считать серого плаща, какой носят иезуиты, но грудь была обнажена, как грудь женщины. Священники все были готовы к воскресной службе, но только один священник был в таком же плаще, как у меня. Все присутствующие думали, что поднимут что-нибудь, как это было принято на Воскресенской службе, но ничего не было поднято, кроме того, что ко мне подошел священник и окропил меня чистой водой.Все присутствующие смеялись, что я одет в такой плащ. Я хотел прикрыть грудь, но, несмотря на все мои старания, она оставалась обнаженной.
Священники все были готовы к воскресной службе, но только один священник был в таком же плаще, как у меня. Все присутствующие думали, что поднимут что-нибудь, как это было принято на Воскресенской службе, но ничего не было поднято, кроме того, что ко мне подошел священник и окропил меня чистой водой.Все присутствующие смеялись, что я одет в такой плащ. Я хотел прикрыть грудь, но, несмотря на все мои старания, она оставалась обнаженной.
791. Во сне видела очень старую женщину, 1500 лет. Ее волосы были белыми, как снег; она принесла мне 2 серебряных пояса и валашскую колбасу. Я купил один у нее и украл другой.
793. Во сне я увидел, что хожу в большую церковь с большим окном, в которой нет ни алтаря, ни картин.Стены были покрыты серебром. Там сидело много польских панов, ели и пили. Они попросили меня поесть с ними, но я сказал, что я слаб и не могу есть. Более того, я еще не слушал мессу. Я пошел в ризницу, и ризница тоже была прекрасна. Я видел, как священник сбросил свою ризу и надел другую. Он пошел молиться с воздетыми руками, но без чаши, войдя в некую комнату, перед которой висела завеса из серебряной материи. Я последовал за ним и увидел человека, лежащего на земле.Он был около 10 локтей в длину и валялся в грязи, но священник помолился ему. Я пошел к тем господам и сказал им: пойдемте, я покажу вам вкусную комедию, как человек валяется в грязи и священник молится ему. Но я был одет в длинную польскую zupania [нижняя одежда польского народного костюма] и подпоясан драгоценным персидским поясом, концы которого были очень драгоценны; и я обмотался несколько раз этим ремнем, но все равно его конец волочился по земле. Сам Господь истолковал: Мне приготовлена какая-то новая дорога.
Он пошел молиться с воздетыми руками, но без чаши, войдя в некую комнату, перед которой висела завеса из серебряной материи. Я последовал за ним и увидел человека, лежащего на земле.Он был около 10 локтей в длину и валялся в грязи, но священник помолился ему. Я пошел к тем господам и сказал им: пойдемте, я покажу вам вкусную комедию, как человек валяется в грязи и священник молится ему. Но я был одет в длинную польскую zupania [нижняя одежда польского народного костюма] и подпоясан драгоценным персидским поясом, концы которого были очень драгоценны; и я обмотался несколько раз этим ремнем, но все равно его конец волочился по земле. Сам Господь истолковал: Мне приготовлена какая-то новая дорога.
804. Господь видел сон 14 июня 1784 года: Пришли ко мне две женщины и один мужчина ростом 6 локтей. Они были очень красивы и сказали мне: «Мы слышали в том месте, где мы живем, что ваши люди покинули вас и что вы не хотите посылать их ни с какой миссией. Мы мертвы уже несколько тысяч лет, и мы много работали, и до сих пор у нас нет покоя. Мы просим вас, пришлите нас. Мы искренне пойдем на вашу миссию. Я ответил им, я уже сказал, что никому не открою, и никого не приведу, и с поручением не пошлю.Меня спрашивали: «Но уже пошёл сигнал, что в мире прольётся много крови, и мы хотим пойти и спасти многих; только ты благословляешь нас на путь. — Я простак, — ответил я, — и благословлять не умею. Они спросили меня: «Но ты благословляешь свой народ?» Я ответил, что могу сказать только это слово: Авраам, Исаак, Иаков и этот стих: Ангел, который избавил меня 1062 & & … Они сказали мне: у нас есть здесь книга, в которой стоят благословения. Благослови нас этой книгой; мы только просим вас благословить нас вслух.Мне дали книгу, написанную крупными еврейскими буквами без точек. Они склонили головы, и я, подняв руки над их головами, благословил их. Там были красивые слова, но я не помню больше двух слов, которые были в конце: Du Jankiew , То есть Джейкоб.
Мы просим вас, пришлите нас. Мы искренне пойдем на вашу миссию. Я ответил им, я уже сказал, что никому не открою, и никого не приведу, и с поручением не пошлю.Меня спрашивали: «Но уже пошёл сигнал, что в мире прольётся много крови, и мы хотим пойти и спасти многих; только ты благословляешь нас на путь. — Я простак, — ответил я, — и благословлять не умею. Они спросили меня: «Но ты благословляешь свой народ?» Я ответил, что могу сказать только это слово: Авраам, Исаак, Иаков и этот стих: Ангел, который избавил меня 1062 & & … Они сказали мне: у нас есть здесь книга, в которой стоят благословения. Благослови нас этой книгой; мы только просим вас благословить нас вслух.Мне дали книгу, написанную крупными еврейскими буквами без точек. Они склонили головы, и я, подняв руки над их головами, благословил их. Там были красивые слова, но я не помню больше двух слов, которые были в конце: Du Jankiew , То есть Джейкоб.
852. Ее Высочество [дочь его Ева] видела сон 5 июля 1784 года: Я видела маленького ребенка в своей комнате; вошел один негр с рогами на голове. Я спросил его: что тебе здесь нужно? Он ответил: я пришел забрать этого ребенка из твоего дома.Я не отдам тебе этого ребенка, сказал я. Он сказал: если ты не отдашь его мне добровольно, то я возьму его силой. Я спросил его, кто он может быть? Он ответил: «Сначала я возьму ребенка, а потом скажу, кто я». Он силой взял ребенка под одну руку, а другой подхватил ту француженку, которая была со мной. Я снова спросил его: кто ты? Он ответил: я худший дьявол из всех дьяволов. Француженка начала громко кричать и просить, чтобы я спас ее от него, но он не послушал ее и ушел с ней.Сразу же в моей комнате начался сильный пожар, который я пытался потушить, но не смог. Господь пришел, чтобы потушить его, на что поднялся большой крик, что в комнате Господа он ужасно горит.
Я спросил его: что тебе здесь нужно? Он ответил: я пришел забрать этого ребенка из твоего дома.Я не отдам тебе этого ребенка, сказал я. Он сказал: если ты не отдашь его мне добровольно, то я возьму его силой. Я спросил его, кто он может быть? Он ответил: «Сначала я возьму ребенка, а потом скажу, кто я». Он силой взял ребенка под одну руку, а другой подхватил ту француженку, которая была со мной. Я снова спросил его: кто ты? Он ответил: я худший дьявол из всех дьяволов. Француженка начала громко кричать и просить, чтобы я спас ее от него, но он не послушал ее и ушел с ней.Сразу же в моей комнате начался сильный пожар, который я пытался потушить, но не смог. Господь пришел, чтобы потушить его, на что поднялся большой крик, что в комнате Господа он ужасно горит.
868. Было бы лучше для тебя, если бы тебя научили премудростям колдовства; вы бы многое знали.
[Первоначально написано для включения, но никогда не публиковалось в Rothenberg & Robinson, Poems for the Millennium , том 3, с биографическими отрывками, адаптированными из H. Л. в Большая еврейская книга (также известная как Изгнанник в Слове ). Джейкоб Франк, еврейский мессия восемнадцатого века, был одним из длинной цепи Мессий со времен Иисуса и до него. См. также H. Lenowitz, The Jewish Messiahs: From the Galilee to Crown Heights , Oxford University Press, 1998. Полное сообщение будет включено как есть в Rothenberg & Bloomberg-Rissman, Barbaric Vast & Wild: An Assemblage of Outside. и подземная поэзия от истоков до наших дней .]
Л. в Большая еврейская книга (также известная как Изгнанник в Слове ). Джейкоб Франк, еврейский мессия восемнадцатого века, был одним из длинной цепи Мессий со времен Иисуса и до него. См. также H. Lenowitz, The Jewish Messiahs: From the Galilee to Crown Heights , Oxford University Press, 1998. Полное сообщение будет включено как есть в Rothenberg & Bloomberg-Rissman, Barbaric Vast & Wild: An Assemblage of Outside. и подземная поэзия от истоков до наших дней .]
15 августа 2014 г.
Извлечение из памяти у мышей и людей
Abstract
Извлечение, т. е. использование полученной информации, до недавнего времени было в основном terra incognita в нейробиологии памяти из-за нехватки методов исследования с пространственно-временным разрешением, необходимым для выявления и анализа быстрой реактивации или реконструкция сложных воспоминаний в мозгу млекопитающих. Развитие новых парадигм, модельных систем и новых инструментов в молекулярной генетике, электрофизиологии, оптогенетике, микроскопии in situ и функциональной визуализации в последние годы заметно способствовало нашей способности исследовать механизмы извлечения мозга. Мы рассматриваем отдельные достижения в изучении явного поиска в мозге грызунов и человека. Возникающая картина заключается в том, что поиск включает в себя скоординированное быстрое взаимодействие разреженных и распределенных сетей кортико-гиппокампа и неокортекса, которые могут позволить перестановочное связывание репрезентативных элементов для получения специфических репрезентаций. Эти репрезентации в значительной степени обусловлены паттернами деятельности, сформированными во время кодирования, но они податливы, подвержены влиянию времени и взаимодействию существующей памяти с новой информацией.
Мы рассматриваем отдельные достижения в изучении явного поиска в мозге грызунов и человека. Возникающая картина заключается в том, что поиск включает в себя скоординированное быстрое взаимодействие разреженных и распределенных сетей кортико-гиппокампа и неокортекса, которые могут позволить перестановочное связывание репрезентативных элементов для получения специфических репрезентаций. Эти репрезентации в значительной степени обусловлены паттернами деятельности, сформированными во время кодирования, но они податливы, подвержены влиянию времени и взаимодействию существующей памяти с новой информацией.
Воспроизведение – это использование усвоенной информации, вызванное сенсорными или внутренними сигналами. В простом модифицированном рефлекторном поведении это относится к считыванию постопытного изменения поведения, вызванного опытом, и лежащей в его основе синаптической эффективности (Кандел и Шварц, 1982; Байрен и Хокинс, 2015). В воспоминаниях, закодированных и хранящихся в более сложных схемах, таких как распределенные воспоминания в мозге млекопитающих, предполагается, что поиск включает в себя различные процессы, включая выбор, реактивацию или реконструкцию целевого представления и оценку результата (Tulving 1983; Dudai 2002). .Эти последовательные и параллельные процессы могут выполняться за долю секунды (например, Thorpe et al. 1996). Поиск имеет решающее значение для понимания памяти. На самом деле, когда кодирование завершено, память, не извлеченная естественным путем или путем экспериментальных манипуляций, не обнаруживается, поэтому извлечение инграммы или ее части является важной частью доказательства существования конкретной инграммы.
.Эти последовательные и параллельные процессы могут выполняться за долю секунды (например, Thorpe et al. 1996). Поиск имеет решающее значение для понимания памяти. На самом деле, когда кодирование завершено, память, не извлеченная естественным путем или путем экспериментальных манипуляций, не обнаруживается, поэтому извлечение инграммы или ее части является важной частью доказательства существования конкретной инграммы.
Несмотря на центральную важность в изучении памяти и обилие данных и моделей припоминания в экспериментальной психологии человека, до недавнего времени припоминание в сложных нейронных схемах оставалось в основном неизведанной территорией в нейробиологии памяти.Это произошло из-за множества препятствий, в том числе трудностей с отделением извлечения от кодирования, ограниченных знаний о локализации конкретных цепей памяти-кандидатов у людей и животных и отсутствия нейробиологических методов с надлежащим пространственно-временным разрешением, позволяющим отслеживать и манипулировать этими цепями. наблюдать, блокировать, запускать или усиливать поиск. Развитие новых парадигм, модельных систем и новых инструментов в молекулярной генетике, электрофизиологии, оптогенетике, микроскопии in situ и функциональной визуализации в последние годы заметно способствовало нашей способности исследовать поиск и понимать часть его процессов и механизмов из клеточных структур. на поведенческий уровень.В этой работе мы рассмотрим некоторые из этих разработок. Мы начнем с избранных исследований восстановления памяти в мозгу грызунов и перейдем к обсуждению аспектов восстановления эпизодической памяти в человеческом мозгу.
наблюдать, блокировать, запускать или усиливать поиск. Развитие новых парадигм, модельных систем и новых инструментов в молекулярной генетике, электрофизиологии, оптогенетике, микроскопии in situ и функциональной визуализации в последние годы заметно способствовало нашей способности исследовать поиск и понимать часть его процессов и механизмов из клеточных структур. на поведенческий уровень.В этой работе мы рассмотрим некоторые из этих разработок. Мы начнем с избранных исследований восстановления памяти в мозгу грызунов и перейдем к обсуждению аспектов восстановления эпизодической памяти в человеческом мозгу.
ВОССТАНОВЛЕНИЕ ПАМЯТИ В МОЗГЕ ГРЫЗУНОВ
Изучение извлечения памяти в мозге млекопитающих предполагает, что этот процесс включает реактивацию паттернов нейронной активности, связанных с первоначальным опытом, хотя и не обязательно идентичных паттернам активности, которые представляли первоначальный опыт.Следовательно, извлечение рассматривается как реконструктивный, а не репликативный процесс. Эта деятельность, вероятно, будет разреженной и анатомически распределенной, при этом различные области мозга вносят вклад в качество и силу припоминания. Хотя работа человека сосредоточена на более богатой психологии и более сложной нейроанатомии, чем у грызунов, общей темой обоих направлений исследований является поиск последовательных паттернов активации, коррелирующих с поиском, и корреляций между поиском и начальным обучением.Работа с грызунами была недавно подстегнута методами, которые позволяют напрямую активировать распределенные нейронные ансамбли, чтобы проверить их функциональное участие в памяти. Сначала мы сосредоточимся на роли гиппокампальных и корковых цепей в извлечении явных воспоминаний у мышей и крыс.
Эта деятельность, вероятно, будет разреженной и анатомически распределенной, при этом различные области мозга вносят вклад в качество и силу припоминания. Хотя работа человека сосредоточена на более богатой психологии и более сложной нейроанатомии, чем у грызунов, общей темой обоих направлений исследований является поиск последовательных паттернов активации, коррелирующих с поиском, и корреляций между поиском и начальным обучением.Работа с грызунами была недавно подстегнута методами, которые позволяют напрямую активировать распределенные нейронные ансамбли, чтобы проверить их функциональное участие в памяти. Сначала мы сосредоточимся на роли гиппокампальных и корковых цепей в извлечении явных воспоминаний у мышей и крыс.
Паттернированная активность во время поиска
Как обсуждалось в литературе, гиппокамп играет решающую роль в явных формах памяти, которые у грызунов широко исследовались в связи с пространственным обучением (Morris et al.1982). Поразительной особенностью гиппокампа грызунов является идентификация клеток места (O’Keefe and Dostrovsky 1971), нейронов, которые возбуждаются, когда животные входят в определенные места в своей среде (Moser et al. 2015). Это привело к мнению, что гиппокамп формирует когнитивную карту; он кодирует карту пространства, которую можно использовать для распознавания определенных сред, направления движения в среде и определения конкретных целевых областей в среде (O’Keefe 1990). На уровне цепи гиппокамп (и окружающие его структуры, такие как энторинальная и периринальная кора) может выполнять эту функцию, интегрируя мультимодальную сенсорную информацию для формирования уникальной карты каждой конкретной среды, объекта или события.Во время извлечения, когда предоставляется достаточный частичный набор сигналов, вся карта (память) рекрутируется таким образом, который, вероятно, включает активацию нескольких областей коры, координируемых гиппокампом. Первый вопрос, который мы можем задать, заключается в том, насколько похожи модели нейронной активности во время двух попыток припоминания или во время припоминания и начального обучения?
2015). Это привело к мнению, что гиппокамп формирует когнитивную карту; он кодирует карту пространства, которую можно использовать для распознавания определенных сред, направления движения в среде и определения конкретных целевых областей в среде (O’Keefe 1990). На уровне цепи гиппокамп (и окружающие его структуры, такие как энторинальная и периринальная кора) может выполнять эту функцию, интегрируя мультимодальную сенсорную информацию для формирования уникальной карты каждой конкретной среды, объекта или события.Во время извлечения, когда предоставляется достаточный частичный набор сигналов, вся карта (память) рекрутируется таким образом, который, вероятно, включает активацию нескольких областей коры, координируемых гиппокампом. Первый вопрос, который мы можем задать, заключается в том, насколько похожи модели нейронной активности во время двух попыток припоминания или во время припоминания и начального обучения?
Одним из способов оценки нейронной активности является исследование группы генов, непосредственно ранних генов (IEG), экспрессия которых в нейронах зависит от активности (Farivar et al. 2004). Наиболее часто используемые ИЭГ для картирования нейронной активности — это cfos , arc и zif268 . Экспрессия этих генов модулируется различными сигнальными путями вторичных мессенджеров, но в возбуждающих нейронах все они, по-видимому, связаны с нервной активностью (Morgan et al., 1987; Sagar et al., 1988). Ген cfos наиболее широко использовался при анализе поведения грызунов и реагирует на всплеск активности 30 или более потенциалов действия с частотой 10 Гц или выше (Schoenenberger et al.2009). Он использовался во многих поведенческих исследованиях, изучающих различные области мозга, и обычно указывает на активность в областях, согласующуюся с известными электрофизиологическими реакциями на поведение. Хотя использование экспрессии IEG предлагает простой анализ нейронной активности с клеточным разрешением, ему не хватает временного разрешения, возможности оценивать низкие уровни активности, например, одиночные потенциалы действия, и способности определять паттерны активности более чем один раз.
2004). Наиболее часто используемые ИЭГ для картирования нейронной активности — это cfos , arc и zif268 . Экспрессия этих генов модулируется различными сигнальными путями вторичных мессенджеров, но в возбуждающих нейронах все они, по-видимому, связаны с нервной активностью (Morgan et al., 1987; Sagar et al., 1988). Ген cfos наиболее широко использовался при анализе поведения грызунов и реагирует на всплеск активности 30 или более потенциалов действия с частотой 10 Гц или выше (Schoenenberger et al.2009). Он использовался во многих поведенческих исследованиях, изучающих различные области мозга, и обычно указывает на активность в областях, согласующуюся с известными электрофизиологическими реакциями на поведение. Хотя использование экспрессии IEG предлагает простой анализ нейронной активности с клеточным разрешением, ему не хватает временного разрешения, возможности оценивать низкие уровни активности, например, одиночные потенциалы действия, и способности определять паттерны активности более чем один раз. точка, которая является требованием для определения стабильности активности нейронного ансамбля во время обучения и поиска или при нескольких попытках поиска.
точка, которая является требованием для определения стабильности активности нейронного ансамбля во время обучения и поиска или при нескольких попытках поиска.
Проблема определения паттернов активности в двух разных временных точках с помощью IEG была решена с помощью метода, называемого компартментальной флуоресцентной гибридизацией in situ (catFISH) (Guzowski et al. 1999). В этом подходе используется тот факт, что гены, которые активно транскрибируются, будут иметь несплайсированную мРНК в ядре, но относительно небольшое количество процессированного транскрипта в цитоплазме в короткие промежутки времени после индукции. Клетки, которые были активны в прошлом, но в настоящее время молчат, будут иметь мРНК в цитоплазме, но не ядерный транскрипт.Подход был использован с геном arc для тестирования ансамблевой активности в гиппокампе у животных, которые дважды исследуют одинаковую среду (А-А), по сравнению с животными, исследующими две разные среды (А-В). Исследование обнаружило большую реактивацию ансамбля в нейронах СА1 (40%), когда две исследуемые среды были идентичными (группа А-А), по сравнению с тем, когда они были разными (группа А-В, реактивация 15%). Этот результат согласуется с идеей о том, что гиппокамп кодирует представление о месте, которое реактивируется при получении информации.Однако в этой интерпретации есть ряд оговорок. Во-первых, эксперимент на самом деле просто сравнивает модель активности двух сенсорных переживаний, а не извлекает воспоминание. Во-вторых, этот метод позволяет сравнивать активность ансамбля только в двух близко расположенных точках времени (30 минут или меньше) и, таким образом, не указывает на стабильность этих ансамблей в течение длительного времени или в нескольких попытках поиска. Наконец, связь ИЭГ с нейронной активностью является грубой и не позволяет точно оценить паттерны активности или низкие уровни активности, как это может быть достигнуто с помощью электрофизиологической записи.
Этот результат согласуется с идеей о том, что гиппокамп кодирует представление о месте, которое реактивируется при получении информации.Однако в этой интерпретации есть ряд оговорок. Во-первых, эксперимент на самом деле просто сравнивает модель активности двух сенсорных переживаний, а не извлекает воспоминание. Во-вторых, этот метод позволяет сравнивать активность ансамбля только в двух близко расположенных точках времени (30 минут или меньше) и, таким образом, не указывает на стабильность этих ансамблей в течение длительного времени или в нескольких попытках поиска. Наконец, связь ИЭГ с нейронной активностью является грубой и не позволяет точно оценить паттерны активности или низкие уровни активности, как это может быть достигнуто с помощью электрофизиологической записи.
Классические методы регистрации гиппокампа не могут легко обнаруживать большие ансамбли нейронов стабильно в течение длительных периодов времени, что затрудняет их использование в экспериментах, описанных выше. Альтернативным подходом является использование генетически кодируемых репортеров уровней кальция в сочетании с оптическим отображением переходных флуоресцентных сигналов, возникающих при связывании кальция (Akerboom et al. 2012). Этот подход предлагает уровень временного разрешения и чувствительности к потенциалу действия, который является промежуточным между физиологической записью и экспрессией IEG, с тем преимуществом, что нейроны точно идентифицируются анатомически и могут стабильно отображаться в течение длительных периодов времени.Этот подход использовался для одновременной регистрации активности от 500 до 1000 нейронов гиппокампа CA1 в течение 45 дней, когда животные бежали по линейной дорожке в постоянной пространственной среде (Ziv et al. 2013). Как видно из предыдущих электрофизиологических исследований, в любой день нейроны демонстрировали четкие пространственные поля возбуждения, при этом около 20% нейронов соответствовали критериям клеток места. Однако между любыми двумя днями точный ансамбль нейронов, задействованных в каждом сеансе, демонстрировал более низкий уровень стабильности.
Альтернативным подходом является использование генетически кодируемых репортеров уровней кальция в сочетании с оптическим отображением переходных флуоресцентных сигналов, возникающих при связывании кальция (Akerboom et al. 2012). Этот подход предлагает уровень временного разрешения и чувствительности к потенциалу действия, который является промежуточным между физиологической записью и экспрессией IEG, с тем преимуществом, что нейроны точно идентифицируются анатомически и могут стабильно отображаться в течение длительных периодов времени.Этот подход использовался для одновременной регистрации активности от 500 до 1000 нейронов гиппокампа CA1 в течение 45 дней, когда животные бежали по линейной дорожке в постоянной пространственной среде (Ziv et al. 2013). Как видно из предыдущих электрофизиологических исследований, в любой день нейроны демонстрировали четкие пространственные поля возбуждения, при этом около 20% нейронов соответствовали критериям клеток места. Однако между любыми двумя днями точный ансамбль нейронов, задействованных в каждом сеансе, демонстрировал более низкий уровень стабильности. При разнице в 5 дней между сеансами записи было 25% совпадение в идентичности ячеек места, записанных в двух сеансах. Перекрытие упало до 15% при 30-дневном интервале между сеансами записи, при этом только ~3% клеток были активны во всех 10 сеансах записи. Таким образом, хотя каждый день 20% нейронов СА1 были активны как клетки места на треке, большинство этих клеток менялись день ото дня в одинаковых пространственных условиях. Если гиппокамп представляет окружающую среду через активность клеток места, то почему не происходит большей стабильности ансамбля нейронов, активируемых, когда животное повторно подвергается воздействию этой среды?
При разнице в 5 дней между сеансами записи было 25% совпадение в идентичности ячеек места, записанных в двух сеансах. Перекрытие упало до 15% при 30-дневном интервале между сеансами записи, при этом только ~3% клеток были активны во всех 10 сеансах записи. Таким образом, хотя каждый день 20% нейронов СА1 были активны как клетки места на треке, большинство этих клеток менялись день ото дня в одинаковых пространственных условиях. Если гиппокамп представляет окружающую среду через активность клеток места, то почему не происходит большей стабильности ансамбля нейронов, активируемых, когда животное повторно подвергается воздействию этой среды?
Есть несколько возможных объяснений очевидной нестабильности ансамблей гиппокампа в предыдущем эксперименте.Одна из возможностей состоит в том, что изо дня в день существуют тонкие различия в окружающей среде, на которую реагирует животное. Другая возможность заключается в том, что задача (бег за водным вознаграждением) не была достаточно заметной, чтобы создать стабильное представление. Предыдущее исследование на мышах с использованием тетродных записей показало, что стабильность клеток места в течение нескольких дней модулируется значимостью задачи, которую животные должны были выполнять в этой среде (Kentros et al. 2004). То, что животные, не лишенные пищи, собирали пищевые гранулы во время записи, приводило к нестабильности клеток места изо дня в день, в то время как от животных требовалось, чтобы они направлялись в определенное место, чтобы избежать неприятного светового / шумового сигнала. временная стабильность в месте возбуждения клеток.Другая возможность состоит в том, что нейроны СА1 кодируют компонент времени таким образом, что каждый последующий день ансамбль меняется, указывая на то, что, хотя сигналы окружающей среды идентичны, сегодняшнее исследование отличается по времени от предыдущих дней. Это было подтверждено электрофизиологической записью клеток места в течение нескольких часов или дней, где было обнаружено, что многие нейроны СА1 изменили свою скорость возбуждения с течением времени, что согласуется с исследованиями визуализации кальция, описанными выше, хотя нейроны СА3 показали большую независимую от сеанса стабильность в паттерне возбуждения.
Предыдущее исследование на мышах с использованием тетродных записей показало, что стабильность клеток места в течение нескольких дней модулируется значимостью задачи, которую животные должны были выполнять в этой среде (Kentros et al. 2004). То, что животные, не лишенные пищи, собирали пищевые гранулы во время записи, приводило к нестабильности клеток места изо дня в день, в то время как от животных требовалось, чтобы они направлялись в определенное место, чтобы избежать неприятного светового / шумового сигнала. временная стабильность в месте возбуждения клеток.Другая возможность состоит в том, что нейроны СА1 кодируют компонент времени таким образом, что каждый последующий день ансамбль меняется, указывая на то, что, хотя сигналы окружающей среды идентичны, сегодняшнее исследование отличается по времени от предыдущих дней. Это было подтверждено электрофизиологической записью клеток места в течение нескольких часов или дней, где было обнаружено, что многие нейроны СА1 изменили свою скорость возбуждения с течением времени, что согласуется с исследованиями визуализации кальция, описанными выше, хотя нейроны СА3 показали большую независимую от сеанса стабильность в паттерне возбуждения. Манкин и др.2012). Таким образом, область CA3 гиппокампа может содержать карту, специфичную для окружающей среды, а область CA1 добавляет информацию, связанную с конкретным временем исследования окружающей среды. Наконец, возможно, что критический пространственный сигнал содержится в небольшом проценте нейронов, которые демонстрируют постоянное возбуждение между несколькими сеансами записи, или в тонкостях паттернов активности, которые ниже порога разрешения этой техники.
Манкин и др.2012). Таким образом, область CA3 гиппокампа может содержать карту, специфичную для окружающей среды, а область CA1 добавляет информацию, связанную с конкретным временем исследования окружающей среды. Наконец, возможно, что критический пространственный сигнал содержится в небольшом проценте нейронов, которые демонстрируют постоянное возбуждение между несколькими сеансами записи, или в тонкостях паттернов активности, которые ниже порога разрешения этой техники.
Каким бы ни было объяснение, этот эксперимент поднимает важный вопрос в понимании того, как представляются и извлекаются воспоминания.Насколько постоянен паттерн мозговой активности в ответ на два идентичных сенсорных входа или два события воспоминаний? Что такое шум и что такое сигнал в наблюдаемом паттерне нейронной активности? Безусловно, способность мозга постоянно распознавать и узнавать об элементах окружающей среды подразумевает некоторый когерентный сигнал в паттернах нейронной активности, вызванных одной и той же сенсорной стимуляцией, но модели и подходы к пониманию этой информации будут различаться в зависимости от того, является ли сигнал доминирующим или нет. второстепенный компонент сенсорно-вызванной активности.
второстепенный компонент сенсорно-вызванной активности.
Исследование паттернов активности во время обучения и припоминания или во время двух приступов припоминания обеспечивает одно из средств идентификации нейронного сигнала, связанного с воспоминанием. Как мы видели выше, эти эксперименты дают несколько противоречивые данные относительно степени достоверности повторной активации ансамбля во время различных событий поиска. Эти эксперименты также проблематичны с концептуальной точки зрения, поскольку они на самом деле не отличают восстановление памяти от сенсорной обработки или кодирования.Когда животных дважды помещают в одну и ту же среду, они испытывают одни и те же сенсорные сигналы, и любая согласованность в активности нейронного ансамбля может отражать обработку этих сигналов, а не поиск в памяти. Наконец, даже если нейронная активность представляет собой восстановление памяти, данные коррелируют и потребуют прямых манипуляций для проверки функциональной релевантности.
Оптогенетические манипуляции во время поиска
Недавно был разработан один подход для прямого тестирования функции распределенных ансамблей активных нейронов у мышей (Reijmers et al. 2007). В этом подходе используется трансгенная мышь, которая позволяет осуществлять генетическую модификацию нейронов на основе их естественных, вызванных окружающей средой моделей активности в пределах экспериментально контролируемого временного окна. IEGs, обсуждавшиеся выше, являются генами, которые экспрессируются в ответ на нервную активность, и предыдущие исследования показали, что их промоторные элементы ДНК могут придавать эту зависимую от активности экспрессию сцепленным репортерам у трансгенных мышей (Smeyne et al. 1992). Это было использовано для разработки бинарной генетической системы, в которой нейронная активность в данный момент времени могла управлять экспрессией любого интересующего гена (GOI) в активных нейронах (14).В этой системе промотор cfos используется для управления экспрессией трансактиватора тетрациклина (tTA), фактора транскрипции, который можно модулировать антибиотиком доксициклином (Dox), производным тетрациклина. tTA может активировать экспрессию второго гена, когда он связан с промоторным элементом, чувствительным к тетрациклину (TRE).
2007). В этом подходе используется трансгенная мышь, которая позволяет осуществлять генетическую модификацию нейронов на основе их естественных, вызванных окружающей средой моделей активности в пределах экспериментально контролируемого временного окна. IEGs, обсуждавшиеся выше, являются генами, которые экспрессируются в ответ на нервную активность, и предыдущие исследования показали, что их промоторные элементы ДНК могут придавать эту зависимую от активности экспрессию сцепленным репортерам у трансгенных мышей (Smeyne et al. 1992). Это было использовано для разработки бинарной генетической системы, в которой нейронная активность в данный момент времени могла управлять экспрессией любого интересующего гена (GOI) в активных нейронах (14).В этой системе промотор cfos используется для управления экспрессией трансактиватора тетрациклина (tTA), фактора транскрипции, который можно модулировать антибиотиком доксициклином (Dox), производным тетрациклина. tTA может активировать экспрессию второго гена, когда он связан с промоторным элементом, чувствительным к тетрациклину (TRE). У животных, несущих два трансгена, cfos-tTA и TRE-GOI, экспрессия tTA напрямую связана с естественной нейронной активностью за счет повышения и понижения промотора cfos по мере того, как нейроны становятся активными.В присутствии Dox транскрипционная активность tTA блокируется, предотвращая активацию ниже по течению от GOI. Когда Dox удаляется, GOI теперь будет экспрессироваться (через транскрипцию, управляемую tTA) во всех нейронах, которые достаточно активны, чтобы управлять cfos-сцепленным tTA. Это позволяет осуществлять генетическую модификацию нейронов, активных в определенные моменты времени в ответ на определенные стимулы окружающей среды, например, сигналы обучения или поиска.
У животных, несущих два трансгена, cfos-tTA и TRE-GOI, экспрессия tTA напрямую связана с естественной нейронной активностью за счет повышения и понижения промотора cfos по мере того, как нейроны становятся активными.В присутствии Dox транскрипционная активность tTA блокируется, предотвращая активацию ниже по течению от GOI. Когда Dox удаляется, GOI теперь будет экспрессироваться (через транскрипцию, управляемую tTA) во всех нейронах, которые достаточно активны, чтобы управлять cfos-сцепленным tTA. Это позволяет осуществлять генетическую модификацию нейронов, активных в определенные моменты времени в ответ на определенные стимулы окружающей среды, например, сигналы обучения или поиска.
Восстановление памяти, вызванное повторной активацией прямой цепи.( A ) Система для введения генов в нейроны на основе их естественных, сенсорно-вызванных паттернов активности (Reijmers et al. 2007). Промотор cfos управляет трансактиватором тетрациклина (tTA) в ответ на нейронную активность, а затем tTA активирует интересующий ген (GOI), который связан с TRE-промотором. Доксициклин (Dox-желтый оттенок) блокирует транскрипционную активность tTA, обеспечивая временной контроль над временными рамками, в которых нейронная активность управляет GOI. ( B ) Экспериментальный план для демонстрации функциональной значимости распределенных нейронных ансамблей.Нейроны, активируемые во время обучения в парадигме обусловливания страха, генетически помечены каналородопсином (ChR2). Последующее индуцированное светом возбуждение этих ансамблей способно вызвать реакцию страха во втором, эмоционально нейтральном контексте.
Доксициклин (Dox-желтый оттенок) блокирует транскрипционную активность tTA, обеспечивая временной контроль над временными рамками, в которых нейронная активность управляет GOI. ( B ) Экспериментальный план для демонстрации функциональной значимости распределенных нейронных ансамблей.Нейроны, активируемые во время обучения в парадигме обусловливания страха, генетически помечены каналородопсином (ChR2). Последующее индуцированное светом возбуждение этих ансамблей способно вызвать реакцию страха во втором, эмоционально нейтральном контексте.
Этот подход использовался в нескольких недавних публикациях для проверки психологической релевантности распределенных нейронных ансамблей, активируемых во время обучения в контекстной задаче на формирование страха (см. Fanselow and Wassum 2015). При контекстуальном обусловливании страха животные получают удары током по ступням в определенном контексте (кондиционирующем ящике), который содержит мультимодальные сенсорные сигналы (как правило, различные визуальные, тактильные и обонятельные сигналы). Восстановление памяти оценивают по реакции страха, которую демонстрируют животные, возвращаясь в камеру кондиционирования. Это модель эксплицитной памяти, которая чувствительна к поражениям гиппокампа и, по сообщениям ряда групп, демонстрирует временной градиент амнезии, наблюдаемый во многих исследованиях пациентов с гиппокампом у людей с более старыми (консолидированными) контекстными воспоминаниями, нечувствительными к поражению гиппокампа (Kim и Fanselow, 1992; Anagnostaras и др., 1999; Squire и Bayley, 2007). В Лю и соавт. (2012), генетическая маркировка cfos использовалась для введения каналородопсина (ChR2) в нейроны зубчатой извилины (DG), которые активировались во время контекстуального обучения страху.Мышей помещали в один из двух контекстов A или B (боксы для кондиционирования с различными визуальными, тактильными и запаховыми сигналами), а нейроны, которые естественным образом активировались в результате этого исследования окружающей среды, помечали ChR2, чтобы обеспечить их последующую прямую активацию светом.
Восстановление памяти оценивают по реакции страха, которую демонстрируют животные, возвращаясь в камеру кондиционирования. Это модель эксплицитной памяти, которая чувствительна к поражениям гиппокампа и, по сообщениям ряда групп, демонстрирует временной градиент амнезии, наблюдаемый во многих исследованиях пациентов с гиппокампом у людей с более старыми (консолидированными) контекстными воспоминаниями, нечувствительными к поражению гиппокампа (Kim и Fanselow, 1992; Anagnostaras и др., 1999; Squire и Bayley, 2007). В Лю и соавт. (2012), генетическая маркировка cfos использовалась для введения каналородопсина (ChR2) в нейроны зубчатой извилины (DG), которые активировались во время контекстуального обучения страху.Мышей помещали в один из двух контекстов A или B (боксы для кондиционирования с различными визуальными, тактильными и запаховыми сигналами), а нейроны, которые естественным образом активировались в результате этого исследования окружающей среды, помечали ChR2, чтобы обеспечить их последующую прямую активацию светом. Затем обе группы мышей были обработаны страхом в контексте А во время приема Dox, чтобы предотвратить дальнейшее мечение активных ансамблей. Когда мыши, у которых DG-нейроны были помечены в контексте A, также были обусловлены страхом в контексте A, последующая стимуляция нейронов, экспрессирующих ChR2, вызывала реакцию страха в нейтральной среде.Стимуляция тех нейронов, которые были генетически помечены ChR2, в то время как животные исследовали контекст B, не вызывала страха у животных, привыкших к контексту страха A. Это говорит о том, что совокупность нейронов, активированных во время исследования контекста A, способна вызывать восстановление памяти при прямой стимуляции. . То есть мозг представляет собой кондиционирующий ящик через активность ансамбля нейронов, активных в DG во время начального исследования (обучения) и, предположительно, через нижестоящие нейроны в других областях мозга, которые активируются, когда этот ансамбль напрямую стимулируется через ChR2.
Затем обе группы мышей были обработаны страхом в контексте А во время приема Dox, чтобы предотвратить дальнейшее мечение активных ансамблей. Когда мыши, у которых DG-нейроны были помечены в контексте A, также были обусловлены страхом в контексте A, последующая стимуляция нейронов, экспрессирующих ChR2, вызывала реакцию страха в нейтральной среде.Стимуляция тех нейронов, которые были генетически помечены ChR2, в то время как животные исследовали контекст B, не вызывала страха у животных, привыкших к контексту страха A. Это говорит о том, что совокупность нейронов, активированных во время исследования контекста A, способна вызывать восстановление памяти при прямой стимуляции. . То есть мозг представляет собой кондиционирующий ящик через активность ансамбля нейронов, активных в DG во время начального исследования (обучения) и, предположительно, через нижестоящие нейроны в других областях мозга, которые активируются, когда этот ансамбль напрямую стимулируется через ChR2.
Весьма примечательно, что крайне нефизиологическая стимуляция, производимая с помощью ChR2, которая возбуждает все нейроны одновременно и, таким образом, устраняет любые эффекты временной последовательности или координацию с эндогенными ритмами, по-видимому, может производить когерентное внутреннее представление сложной окружающей среды. Для дальнейшего изучения этой идеи был проведен дополнительный эксперимент, чтобы определить, может ли ансамблевая активность в DG служить условным стимулом (CS) при обусловливании страха.В этом случае cfos -активированных нейронов были генетически помечены ChR2, в то время как животные исследовали контекст А, как и в предыдущем исследовании. Однако теперь ансамбль, меченый ChR2, активировался в нейтральном контексте и сочетался с электрошоком. Таким образом, искусственная стимуляция выполняла роль СУ, которая сочеталась с безусловным раздражителем (УЗ) электрошоком. В этом случае у животных развилась реакция страха на контекст А, даже несмотря на то, что они фактически никогда не подвергались аверсивному шоку США в этой среде (Ramirez et al.2013). В совокупности эти результаты подтверждают мнение о том, что активность нейронных ансамблей DG достаточна для представления контекста и служит сигналом для кодирования и извлечения памяти.
Для дальнейшего изучения этой идеи был проведен дополнительный эксперимент, чтобы определить, может ли ансамблевая активность в DG служить условным стимулом (CS) при обусловливании страха.В этом случае cfos -активированных нейронов были генетически помечены ChR2, в то время как животные исследовали контекст А, как и в предыдущем исследовании. Однако теперь ансамбль, меченый ChR2, активировался в нейтральном контексте и сочетался с электрошоком. Таким образом, искусственная стимуляция выполняла роль СУ, которая сочеталась с безусловным раздражителем (УЗ) электрошоком. В этом случае у животных развилась реакция страха на контекст А, даже несмотря на то, что они фактически никогда не подвергались аверсивному шоку США в этой среде (Ramirez et al.2013). В совокупности эти результаты подтверждают мнение о том, что активность нейронных ансамблей DG достаточна для представления контекста и служит сигналом для кодирования и извлечения памяти.
Искусственная стимуляция небольших ансамблей нейронов в области DG гиппокампа позволяет извлекать контекстуальные воспоминания, предположительно, путем активации более распределенных ансамблей нейронов в нижестоящих областях гиппокампа и коры. Учитывая сильно параллельный характер нейронной связи и обработки, возможно, что, хотя этих ансамблей может быть достаточно для поиска, они не являются необходимыми, поскольку другие пути могут компенсировать это.Вопрос о необходимости рассматривался в двух недавних исследованиях с использованием светозависимого протонного насоса архебактерий (ArchT) для гиперполяризации и молчания cfos-меченых нейронных ансамблей в экспериментах, аналогичных описанным выше с ChR2 (Denny et al., 2014; Tanaka et al. 2014). Обе группы исследователей также использовали контекстуальное обусловливание страха и изучили потребность в ансамблях в трех разных областях гиппокампа: DG, CA3 и CA1. Экспрессия ArchT была направлена на нейроны, которые были активны во время обучения в условиях обусловленного контекстом страха, и эти ансамбли впоследствии были подавлены либо в DG, CA3, либо в CA1 во время извлечения памяти.В каждом случае замалчивание нарушало восстановление контекстуальной памяти о страхе. Эти результаты показывают, что в каждой из трех основных субрегионов гиппокампа реактивация ансамбля нейронов, активных во время начального обучения, необходима для последующего извлечения памяти.
Учитывая сильно параллельный характер нейронной связи и обработки, возможно, что, хотя этих ансамблей может быть достаточно для поиска, они не являются необходимыми, поскольку другие пути могут компенсировать это.Вопрос о необходимости рассматривался в двух недавних исследованиях с использованием светозависимого протонного насоса архебактерий (ArchT) для гиперполяризации и молчания cfos-меченых нейронных ансамблей в экспериментах, аналогичных описанным выше с ChR2 (Denny et al., 2014; Tanaka et al. 2014). Обе группы исследователей также использовали контекстуальное обусловливание страха и изучили потребность в ансамблях в трех разных областях гиппокампа: DG, CA3 и CA1. Экспрессия ArchT была направлена на нейроны, которые были активны во время обучения в условиях обусловленного контекстом страха, и эти ансамбли впоследствии были подавлены либо в DG, CA3, либо в CA1 во время извлечения памяти.В каждом случае замалчивание нарушало восстановление контекстуальной памяти о страхе. Эти результаты показывают, что в каждой из трех основных субрегионов гиппокампа реактивация ансамбля нейронов, активных во время начального обучения, необходима для последующего извлечения памяти. Это согласуется с мнением о том, что припоминание включает в себя реконструкцию паттернов мозговой активности, созданных во время начального обучения.
Это согласуется с мнением о том, что припоминание включает в себя реконструкцию паттернов мозговой активности, созданных во время начального обучения.
Извлечение консолидированных воспоминаний
Другим примечательным аспектом памяти и ее извлечения является изменение структуры цепи воспоминаний, зависящих от гиппокампа, с течением времени, первоначально представленное у пациентов с амнезией, таких как H.М. (Сквайр и Деде, 2015). Наблюдение, что вновь сформированные воспоминания требуют гиппокампа для извлечения, но спустя недели (у грызунов) или месяцы или годы (у людей) гиппокамп, по-видимому, становится необязательным, по крайней мере, для некоторых форм эксплицитной памяти, предполагает, что имеет место анатомическое изменение. в памяти со временем. Какова природа этой консолидации памяти на основе цепей («консолидация систем») (Squire et al. 2015)? Каково анатомическое местонахождение зависимых от гиппокампа новых воспоминаний и независимых от гиппокампа старых, консолидированных воспоминаний? Какова роль гиппокампа и коры в кодировании и извлечении новых и старых воспоминаний? Эти вопросы также всплывут ниже при обсуждении эпизодического поиска у людей. Текущий взгляд на этот процесс постулирует, что во время первоначального кодирования и извлечения эксплицитной памяти гиппокамп играет решающую роль, например, кодируя индекс или карту, которая помогает задействовать соответствующие области коры и определенные нейронные ансамбли для воспроизведения. Со временем эти корковые ансамбли могут стать независимыми от гиппокампа, возможно, за счет усиления их связности за счет повторения соответствующих паттернов активности после обучения (Wilson and McNaughton, 1994), так что с новыми усиленными корковыми связями память или, по крайней мере, некоторые обработанные версии его можно получить независимо от гиппокампа.
Текущий взгляд на этот процесс постулирует, что во время первоначального кодирования и извлечения эксплицитной памяти гиппокамп играет решающую роль, например, кодируя индекс или карту, которая помогает задействовать соответствующие области коры и определенные нейронные ансамбли для воспроизведения. Со временем эти корковые ансамбли могут стать независимыми от гиппокампа, возможно, за счет усиления их связности за счет повторения соответствующих паттернов активности после обучения (Wilson and McNaughton, 1994), так что с новыми усиленными корковыми связями память или, по крайней мере, некоторые обработанные версии его можно получить независимо от гиппокампа.
Доказательства такого взгляда на консолидированные воспоминания у грызунов получены в результате поражения и инактивации определенных областей мозга, а также в исследованиях нейронной активности, возникающей при извлечении недавних и отдаленных воспоминаний. Одно из предсказаний классической модели консолидации памяти состоит в том, что, хотя гиппокамп играет важную роль в извлечении недавних воспоминаний, области коры вместо этого должны требоваться для извлечения отдаленных воспоминаний или, возможно, недавних и отдаленных воспоминаний. Это было проверено при консолидации трех различных парадигм эксплицитной памяти у грызунов: пространственной памяти, контекстуальной памяти и социальной памяти, основанной на обонянии (Bontempi et al.1999 г.; Франкленд и др. 2004 г.; Франкленд и Бонтемпи, 2005 г.). Во всех трех исследованиях повреждения гиппокампа нарушали недавнее восстановление памяти (через 1 день после тренировки), но сохраняли отдаленное извлечение (30 дней после тренировки), как и ожидалось для зависимых от гиппокампа задач, связанных с консолидацией. Что касается контекстуального страха и пространственных воспоминаний, инактивация передней поясной коры вызывала нарушение извлечения отдаленных воспоминаний, но не влияла на недавние воспоминания, что противоположно тому, что наблюдается при инактивации или поражении гиппокампа (Kim and Fanselow 1992; Anagnostaras et al.1999). Аналогичный результат наблюдался с префронтальной инактивацией в пространственной задаче, в то время как в обонятельной социальной задаче инактивация орбитальной лобной коры нарушала как недавнее, так и отдаленное воспоминание.
Это было проверено при консолидации трех различных парадигм эксплицитной памяти у грызунов: пространственной памяти, контекстуальной памяти и социальной памяти, основанной на обонянии (Bontempi et al.1999 г.; Франкленд и др. 2004 г.; Франкленд и Бонтемпи, 2005 г.). Во всех трех исследованиях повреждения гиппокампа нарушали недавнее восстановление памяти (через 1 день после тренировки), но сохраняли отдаленное извлечение (30 дней после тренировки), как и ожидалось для зависимых от гиппокампа задач, связанных с консолидацией. Что касается контекстуального страха и пространственных воспоминаний, инактивация передней поясной коры вызывала нарушение извлечения отдаленных воспоминаний, но не влияла на недавние воспоминания, что противоположно тому, что наблюдается при инактивации или поражении гиппокампа (Kim and Fanselow 1992; Anagnostaras et al.1999). Аналогичный результат наблюдался с префронтальной инактивацией в пространственной задаче, в то время как в обонятельной социальной задаче инактивация орбитальной лобной коры нарушала как недавнее, так и отдаленное воспоминание.
В дополнение к инактивации и повреждениям для изучения анатомической структуры недавних и отдаленных воспоминаний у грызунов в этих исследованиях также изучались паттерны нейронной активации во время поиска с использованием экспрессии IEG. В каждой поведенческой парадигме активация гиппокампа была выше при извлечении недавних воспоминаний, а не отдаленных.И наоборот, широкий спектр областей коры проявлял повышенную активность во время извлечения отдаленных по сравнению с недавними воспоминаний. Хотя эти исследования подтверждают точку зрения, согласно которой отдаленные воспоминания требуют повышенной роли коры в обработке, они также поднимают некоторые вопросы, касающиеся исходной модели координации гиппокампа корковой активности во время извлечения недавних воспоминаний. Если во время извлечения недавней памяти гиппокамп координирует и рекрутирует корковую активность, то почему эти области часто не требуются для извлечения в этот момент времени и почему корковая активность изменяется с течением времени. Это говорит о том, что со временем в схемах и характере этих воспоминаний могут произойти более фундаментальные изменения. Это очевидно на поведенческом уровне, поскольку исследования на мышах (и людях) показали, что отдаленные воспоминания теряют некоторую специфичность (Wiltgen and Silva, 2007; Winocur et al., 2010; Furman et al., 2012).
Это говорит о том, что со временем в схемах и характере этих воспоминаний могут произойти более фундаментальные изменения. Это очевидно на поведенческом уровне, поскольку исследования на мышах (и людях) показали, что отдаленные воспоминания теряют некоторую специфичность (Wiltgen and Silva, 2007; Winocur et al., 2010; Furman et al., 2012).
Одно недавнее исследование на мышах изучало кортикальное представление эксплицитной памяти с помощью мечения поведенчески активных нейронных ансамблей с помощью ChR2 (Cowansage et al. 2014).В экспериментах, аналогичных описанным выше для DG, экспрессия ChR2 была направлена на нейроны, которые активировались во время контекстного страха. Исследователи исследовали ретросплениальную кору, выходную область коры гиппокампа, которая необходима для кодирования и извлечения как недавней, так и отдаленной контекстуальной памяти (Keene and Bucci 2008a,b; см. также исследования на людях ниже). Они обнаружили, что искусственная стимуляция ансамбля, активируемого обучением (через ChR2), вызывала реакцию замирания, предполагая, что она инициировала событие припоминания, подобное тому, что наблюдалось в исследованиях гиппокампа. Это показывает контекстуальное представление, способное производить поисковые формы в коре, в дополнение к гиппокампу, во время обучения. Что еще более удивительно, они обнаружили, что искусственная стимуляция этих ансамблей ретроспленальной коры вызывала воспоминания о страхе, даже когда гиппокамп был фармакологически заглушен через 1 день после тренировки, когда гиппокамп все еще необходим для естественного извлечения контекстуальных воспоминаний. Эти результаты показывают не только то, что корковое представление для контекстной памяти формируется во время обучения, но и нормальную потребность гиппокампа в воспоминании можно обойти путем прямой активации этого представления.Этот вывод согласуется с представлением о гиппокампе как о карте или указателе, который задействует соответствующие корковые цепи во время поиска в памяти, но не хранит напрямую необходимый компонент самого консолидированного представления.
Это показывает контекстуальное представление, способное производить поисковые формы в коре, в дополнение к гиппокампу, во время обучения. Что еще более удивительно, они обнаружили, что искусственная стимуляция этих ансамблей ретроспленальной коры вызывала воспоминания о страхе, даже когда гиппокамп был фармакологически заглушен через 1 день после тренировки, когда гиппокамп все еще необходим для естественного извлечения контекстуальных воспоминаний. Эти результаты показывают не только то, что корковое представление для контекстной памяти формируется во время обучения, но и нормальную потребность гиппокампа в воспоминании можно обойти путем прямой активации этого представления.Этот вывод согласуется с представлением о гиппокампе как о карте или указателе, который задействует соответствующие корковые цепи во время поиска в памяти, но не хранит напрямую необходимый компонент самого консолидированного представления.
Эксперименты с использованием ChR2 для реактивации нейронных ансамблей, которые естественным образом активировались во время обучения, дают важную информацию о том, как мозг хранит и представляет сложную информацию о внешнем мире. Однако может быть неправильно характеризовать эти нейроны как «энграммы» или нейроны, содержащие инграммы, как это иногда предлагалось.Термин «энграмма» относится к физическим изменениям в мозгу, лежащим в основе памяти (Semon 1904; Lashley 1950). Если мы возьмем контекстное обусловливание в качестве примера, поле обусловливания изначально нейтрально, и воздействие вызывает исследовательское поведение. В сочетании с электрошоком та же самая камера теперь вызывает у животных реакцию страха. Инграммой для этой памяти будут участки в мозгу, которые направляют ту же сенсорную информацию (подсказки в коробке) для обработки таким образом, чтобы вызвать страх, а не исследовательское поведение, например, направляя информацию для активации миндалевидного тела. .Можно ожидать, что стимуляция ансамблей ChR2 в любой точке пути обработки сенсорной информации контекста представляет контекст и, следовательно, заменяет его, но не является на самом деле критическим участком пластичности, необходимым для создания цепей обработки страха.
Однако может быть неправильно характеризовать эти нейроны как «энграммы» или нейроны, содержащие инграммы, как это иногда предлагалось.Термин «энграмма» относится к физическим изменениям в мозгу, лежащим в основе памяти (Semon 1904; Lashley 1950). Если мы возьмем контекстное обусловливание в качестве примера, поле обусловливания изначально нейтрально, и воздействие вызывает исследовательское поведение. В сочетании с электрошоком та же самая камера теперь вызывает у животных реакцию страха. Инграммой для этой памяти будут участки в мозгу, которые направляют ту же сенсорную информацию (подсказки в коробке) для обработки таким образом, чтобы вызвать страх, а не исследовательское поведение, например, направляя информацию для активации миндалевидного тела. .Можно ожидать, что стимуляция ансамблей ChR2 в любой точке пути обработки сенсорной информации контекста представляет контекст и, следовательно, заменяет его, но не является на самом деле критическим участком пластичности, необходимым для создания цепей обработки страха. инграмма). Например, если бы сетчатку можно было искусственно стимулировать точно таким же образом, как когда животное исследует контекст, то можно было бы ожидать, что она вызовет реакцию страха у обусловленных животных таким же образом, как в экспериментах, описанных выше, но без переноса «инграммы». «за эту память.
инграмма). Например, если бы сетчатку можно было искусственно стимулировать точно таким же образом, как когда животное исследует контекст, то можно было бы ожидать, что она вызовет реакцию страха у обусловленных животных таким же образом, как в экспериментах, описанных выше, но без переноса «инграммы». «за эту память.
Изучение памяти беспозвоночных создает прочную концептуальную основу для понимания обработки информации нервной системой. Сенсорный ввод преобразуется в различные двигательные/поведенческие результаты на основе опыта и посредством пластических изменений в определенных узлах сети обработки. Мозг млекопитающих вводит увеличение количества нейронов между входными и выходными узлами на 4 (мыши) и 7 (человека) порядков, но глубокое понимание памяти и поиска млекопитающих по-прежнему требует определения того, как эта информация проходит через и представлен в нервной системе и определенных участках, которые изменяются с опытом, чтобы произвести извлекаемую память.Мы представили некоторые методы, используемые в моделях мышей и крыс, и результаты, которые предлагают основу для исследования этой обработки на тонком уровне, ориентированном на определенные нейронные ансамбли. Дополнительная сложность мозга млекопитающих позволила увеличить специализацию областей для различных задач обработки, а память/воспроизведение, вероятно, будет включать скоординированное взаимодействие более разреженных и распределенных сетей, чем в более простых системах. Работа с людьми, включающая функциональную визуализацию мозга и, в последнее время, электрофизиологию, также начинает определять некоторые принципы этих сетей обработки данных.Эти методы позволяют оценить активность всего мозга во время кодирования и извлечения и обеспечивают представление о взаимодействиях и функциональных различиях различных областей во время обработки.
Дополнительная сложность мозга млекопитающих позволила увеличить специализацию областей для различных задач обработки, а память/воспроизведение, вероятно, будет включать скоординированное взаимодействие более разреженных и распределенных сетей, чем в более простых системах. Работа с людьми, включающая функциональную визуализацию мозга и, в последнее время, электрофизиологию, также начинает определять некоторые принципы этих сетей обработки данных.Эти методы позволяют оценить активность всего мозга во время кодирования и извлечения и обеспечивают представление о взаимодействиях и функциональных различиях различных областей во время обработки.
ПОИСК ПАМЯТИ В ЧЕЛОВЕЧЕСКОМ МОЗГЕ
Наше обсуждение восстановления человеческой памяти будет сосредоточено на декларативной и особенно эпизодической памяти (Squire and Dede 2015). Мы начнем с краткого обзора аспектов богатого феноменологического анализа извлечения информации из памяти человека, который заложил основы многих современных исследований мозговых субстратов и процессов извлечения информации. Затем мы рассмотрим современные знания о цепях человеческого мозга, которые служат поиску, и закончим кратким описанием функциональных моделей, которые лежат в основе многих исследований мозга долговременного поиска памяти.
Затем мы рассмотрим современные знания о цепях человеческого мозга, которые служат поиску, и закончим кратким описанием функциональных моделей, которые лежат в основе многих исследований мозга долговременного поиска памяти.
Феноменологический анализ процессов извлечения информации у человека
Современные исследования в области нейробиологии восстановления человеческой памяти в значительной степени опираются на богатый массив исследований в области экспериментальной и когнитивной психологии, которые процветали уже полвека назад. Это исследование дало классификации, модели и вопросы, которые находятся в авангарде изучения мозга и поведенческих механизмов поиска.Мы упомянем лишь несколько примеров, а затем перейдем к описанию их отражения в исследованиях мозга.
Основное различие в декларативном поиске человеком заключается в воспроизведении и распознавании. В то время как припоминание — это повторная активация или реконструкция внутреннего представления целевого объекта в отсутствие этого объекта, вызванная неявными или явными сигналами, узнавание — это суждение о предыдущем появлении в присутствии хотя бы части целевого объекта. Таким образом, это различие относится как к тесту, используемому для исследования памяти (т.т. е., в присутствии или в отсутствие целевого элемента) и постулируемой когнитивной основе производительности памяти, измеряемой тестом. Было предложено, чтобы припоминание и распознавание включали несколько процессов, а сами эти процессы были дополнительно разобщены. Короче говоря, первоначально некоторые предполагали, что припоминание состоит из двух основных фаз: «фазы генерации», за которой следует фаза узнавания (Bahrick, 1970; Kintsch, 1970; Anderson and Bower, 1972). Точно так же модели узнавания с «двойным процессом» утверждают, что суждения об узнавании могут быть основаны на двух различных типах памяти: знакомстве и воспоминании (James 1890; Mandler 1980; Jacoby and Dallas 1981; Yonelinas 1994; Diana et al.2006 г.; Восс и Паллер, 2010). Многочисленные экспериментальные парадигмы использовались для разделения этих двух процессов (Yonelinas 2002), исследуя либо субъективное чувство воспоминания (участники указывают, вспоминают ли они предмет, либо находят его знакомым при отсутствии конкретного воспоминания [Tulving 1985]), либо объективная способность вспомнить дополнительные аспекты изучаемого события, такие как контекст, в котором оно было изучено («исходная память»), или элемент, связанный с зондом во время обучения.
Таким образом, это различие относится как к тесту, используемому для исследования памяти (т.т. е., в присутствии или в отсутствие целевого элемента) и постулируемой когнитивной основе производительности памяти, измеряемой тестом. Было предложено, чтобы припоминание и распознавание включали несколько процессов, а сами эти процессы были дополнительно разобщены. Короче говоря, первоначально некоторые предполагали, что припоминание состоит из двух основных фаз: «фазы генерации», за которой следует фаза узнавания (Bahrick, 1970; Kintsch, 1970; Anderson and Bower, 1972). Точно так же модели узнавания с «двойным процессом» утверждают, что суждения об узнавании могут быть основаны на двух различных типах памяти: знакомстве и воспоминании (James 1890; Mandler 1980; Jacoby and Dallas 1981; Yonelinas 1994; Diana et al.2006 г.; Восс и Паллер, 2010). Многочисленные экспериментальные парадигмы использовались для разделения этих двух процессов (Yonelinas 2002), исследуя либо субъективное чувство воспоминания (участники указывают, вспоминают ли они предмет, либо находят его знакомым при отсутствии конкретного воспоминания [Tulving 1985]), либо объективная способность вспомнить дополнительные аспекты изучаемого события, такие как контекст, в котором оно было изучено («исходная память»), или элемент, связанный с зондом во время обучения.
Хотя многие исследователи согласны с существованием двух различных процессов распознавания (но см. Shimamura 2010, модель с одним процессом), остается несколько спорных споров. Первый из них относится к природе сигнала воспоминаний и к тому, как воспоминание и знакомство в конечном итоге способствуют оценке узнавания. Два ведущих класса моделей, каждая из которых основана как на поведенческих, так и на функциональных данных нейровизуализации, различаются в первую очередь своим взглядом на воспоминание либо как на пороговый процесс, либо как на непрерывную переменную.Согласно двухпроцессной модели обнаружения сигналов (Йонелинас, 1994, 2002; Паркс и Йонелинас, 2007), знакомство и воспоминание представляют собой два независимых процесса, инициируемых параллельно. Считается, что знакомство отражает непрерывную меру силы памяти, которую лучше всего моделировать как процесс обнаружения сигналов. И наоборот, припоминание считается пороговым процессом, при котором будут вспоминаться только те элементы, которые превышают определенный порог, что приводит к относительно высоконадежным ответам (Yonelinas et al. 2010).
2010).
Согласно альтернативному подходу, воспоминание также является непрерывной мерой, которая лучше всего моделируется процессом обнаружения сигнала, а суждения о узнавании основаны на агрегированной переменной силы памяти (Rotello et al. 2004; Wixted 2007; Mickes et al. , 2009; Викстед и Микес, 2010). Модель непрерывного двойного процесса (Wixted and Mickes, 2010) утверждает, что в процессе узнавания воспоминание и знакомство вызывают отдельную внутреннюю меру вдоль оси силы памяти.Критерий принятия решения для идентификации предмета как распознанного может быть основан либо на одной из этих осей, либо на агрегированной оси памяти-силы, которая учитывает как чувство припоминания, так и чувство знакомства, в зависимости от поставленной задачи. Различные подходы начинают сходиться, с предположениями, что воспоминание может оцениваться на основе количества воспоминаемой информации (Rugg et al. 2012), но все же подвергаться пороговому процессу (Yonelinas and Jacoby 2012).
Еще один спорный вопрос заключается в том, являются ли тесты, обычно используемые для разграничения воспоминаний и знакомства (т. например, тест «помнить/знать» [Tulving 1985]) действительно отделяют эти два процесса (например, Perfect 1996; Yonelinas 2002; Parks and Yonelinas 2007; Yonelinas et al. 2010), или же они на самом деле только отделяют сильные воспоминания от более слабых (Дональдсон, 1996; Данн, 2004; Вайс и др., 2008; Викстед и др., 2010). Наконец, что не менее важно, в контексте настоящего обсуждения дебаты касаются роли отдельных областей мозга, прежде всего медиальной височной доли (MTL), в процессах воспоминания/знакомства (см. ниже).
например, тест «помнить/знать» [Tulving 1985]) действительно отделяют эти два процесса (например, Perfect 1996; Yonelinas 2002; Parks and Yonelinas 2007; Yonelinas et al. 2010), или же они на самом деле только отделяют сильные воспоминания от более слабых (Дональдсон, 1996; Данн, 2004; Вайс и др., 2008; Викстед и др., 2010). Наконец, что не менее важно, в контексте настоящего обсуждения дебаты касаются роли отдельных областей мозга, прежде всего медиальной височной доли (MTL), в процессах воспоминания/знакомства (см. ниже).
В качестве примера мы выбрали дискуссию о воспоминании/знакомстве, так как в настоящее время это одна из самых горячих тем в исследованиях извлечения эпизодической памяти (Voss and Paller 2010). Однако следует отметить, что это далеко не все исследования феноменологии и, в конечном счете, потенциальных мозговых механизмов извлечения человеческой памяти. Проводится дополнительное различие между содержанием поиска и входом в состояние, которое делает возможным поиск («режим поиска» [Tulving 1983], см. Rugg and Wilding 2000, для более детального разделения) или, в более общем смысле, между различными типами специфических для элемента и инвариантных для элемента процессов при поиске (т.г., Найберг и др. 1995 год; Бакнер и др. 1998 год; Кёлер и др. 1998 год; Доббинс и Вагнер, 2005 г.; Дуарте и др. 2011 г.; Бергстрём и др. 2013). Кроме того, было показано, что разные типы воспоминаний включают разные процессы извлечения, например, при сравнении автобиографических и лабораторных воспоминаний (Cabeza et al., 2004; Svoboda et al., 2006; McDermott et al., 2009), поле и точка зрения наблюдателя. (Eich et al. 2009), объективные и субъективные измерения воспоминаний (см. Spaniol et al. 2009 для метаанализа) и эмоциональные и нейтральные воспоминания (Maratos and Rugg 2001; LaBar and Cabeza 2006; Buchanan 2007).Хотя это ограниченное обсуждение не позволяет провести всесторонний обзор мелкозернистого анализа феноменологии человеческого поиска, важно иметь в виду, что богатая дифференциация процесса поиска проявляется в различной активности мозга во время поиска.
Rugg and Wilding 2000, для более детального разделения) или, в более общем смысле, между различными типами специфических для элемента и инвариантных для элемента процессов при поиске (т.г., Найберг и др. 1995 год; Бакнер и др. 1998 год; Кёлер и др. 1998 год; Доббинс и Вагнер, 2005 г.; Дуарте и др. 2011 г.; Бергстрём и др. 2013). Кроме того, было показано, что разные типы воспоминаний включают разные процессы извлечения, например, при сравнении автобиографических и лабораторных воспоминаний (Cabeza et al., 2004; Svoboda et al., 2006; McDermott et al., 2009), поле и точка зрения наблюдателя. (Eich et al. 2009), объективные и субъективные измерения воспоминаний (см. Spaniol et al. 2009 для метаанализа) и эмоциональные и нейтральные воспоминания (Maratos and Rugg 2001; LaBar and Cabeza 2006; Buchanan 2007).Хотя это ограниченное обсуждение не позволяет провести всесторонний обзор мелкозернистого анализа феноменологии человеческого поиска, важно иметь в виду, что богатая дифференциация процесса поиска проявляется в различной активности мозга во время поиска. подчеркивая, что поиск не может быть исследован как единая конструкция.
подчеркивая, что поиск не может быть исследован как единая конструкция.
Гиппокамп и MTL
Большое внимание в изучении эпизодического поиска было уделено роли MTL (Squire and Dede 2015), в частности, дифференциации между гиппокампом и окружающей корой.Поскольку очаговое повреждение свода и сосцевидных тел приводит к нарушению работоспособности, напоминающему повреждение гиппокампа (Dusoir et al., 1990; Tsivilis et al., 2008; Rudebeck et al., 2009; Vann et al., 2009b), было высказано предположение, что изучение памяти на основе гиппокампа должно быть расширено, чтобы включить свод и сосцевидные тела как часть «расширенной системы гиппокампа» (Aggleton and Brown 1999).
Один из подходов к дифференциации вклада гиппокампа и коры MTL в припоминание основан на вышеупомянутом различии воспоминание/знакомство.В нескольких исследованиях рассматривался вопрос о том, участвует ли гиппокамп в воспоминании исключительно, или же он поддерживает как знакомство, так и воспоминание (Brown and Aggleton, 2001; Eichenbaum et al. , 2007; Skinner and Fernandes, 2007; Squire et al., 2007; Wixted and Squire). 2011; Рагг и др., 2012; Рагг и Вилберг, 2013). Обе точки зрения основаны на результатах исследований пациентов с повреждением MTL в сочетании с результатами функциональной нейровизуализации у здоровых людей.
, 2007; Skinner and Fernandes, 2007; Squire et al., 2007; Wixted and Squire). 2011; Рагг и др., 2012; Рагг и Вилберг, 2013). Обе точки зрения основаны на результатах исследований пациентов с повреждением MTL в сочетании с результатами функциональной нейровизуализации у здоровых людей.
В то время как во многих исследованиях сообщается о непропорциональном влиянии повреждения гиппокампа на припоминание и ассоциативную память по сравнению со знакомостью (Huppert and Piercy 1978; Vargha-Khadem et al.1997 год; Холдсток и др. 2002 г.; Йонелинас и др. 2002 г.; Джованелло и др. 2003 г.; Мэйс и др. 2004 г.; Аглтон и др. 2005), другие отчеты показывают, что повреждение гиппокампа влияет на знакомство и память в аналогичной степени (Manns and Squire 1999; Stark et al. 2002; Manns et al. 2003; Cipolotti et al. 2006; Wais et al. 2006; Jeneson et al. , 2010; Кирван и др., 2010; Сонг и др., 2011). Интересно, что у пациента со значительным повреждением околоносовых пазух, не затрагивающим гиппокамп, было нарушено знакомство и сохранена память (Bowles et al. 2007). Такие исследования указывают на причинно-следственную роль областей MTL в различных процессах, но не допускают диссоциации между кодированием, хранением и поиском, а также не обязательно отражают процессы памяти в здоровом мозге.
2007). Такие исследования указывают на причинно-следственную роль областей MTL в различных процессах, но не допускают диссоциации между кодированием, хранением и поиском, а также не обязательно отражают процессы памяти в здоровом мозге.
И наоборот, исследования функциональной магнитно-резонансной томографии (фМРТ) не позволяют продемонстрировать причинно-следственную связь, но позволяют целенаправленно исследовать корреляцию с различными стадиями памяти. Однако даже в исследованиях фМРТ, посвященных процессам поиска, случайное кодирование во время задач поиска может препятствовать способности различать процессы кодирования и поиска (Buckner et al.2001 г.; Старк и Окадо 2003; Ким 2013; но см. Staresina et al. 2012б; Бен-Яков и др. 2014 для временных диссоциаций между кодированием и поиском). Как и в случае с пациентами, исследования фМРТ привели к расходящимся результатам в отношении участия гиппокампа в воспоминании. Одна серия исследований показала, что гиппокамп служит воспоминанию, но не знакомству с меморандумами, такими как слова или изображения (Eldridge et al. , 2000, 2005; Weis et al., 2004; Wheeler and Buckner, 2004; Yonelinas et al., 2005; Montaldi et al. др.2006 г.; Диана и др. 2007, 2010; Кон и др. 2009 г.; Форд и др. 2010 г.; Рагг и др. 2012 г.; Ю и др. 2012). В частности, было обнаружено, что гиппокамп более сильно реагирует на слова или изображения, о которых сообщается, что они «вспоминаются» (Элдридж и др., 2000, 2005; Уилер и Бакнер, 2004; Йонелинас и др., 2005; Монтальди и др., 2006), в правильных выражениях. по сравнению с неправильным поиском контекста кодирования (Weis et al. 2004; Tendolkar et al. 2008; Duarte et al. 2011; см. Staresina et al. 2012b для связанного исследования внутричерепной электроэнцефалографии [iEEG]) и в ассоциативном по сравнению с неассоциативным распознаванием (Кирван и Старк, 2004) или распознавание сложных слов (Форд и др.2010). Недавние исследования предполагают, что активность гиппокампа не связана с субъективным чувством воспоминания, а модулируется объемом фактически извлеченной контекстуальной информации (Rugg et al.
, 2000, 2005; Weis et al., 2004; Wheeler and Buckner, 2004; Yonelinas et al., 2005; Montaldi et al. др.2006 г.; Диана и др. 2007, 2010; Кон и др. 2009 г.; Форд и др. 2010 г.; Рагг и др. 2012 г.; Ю и др. 2012). В частности, было обнаружено, что гиппокамп более сильно реагирует на слова или изображения, о которых сообщается, что они «вспоминаются» (Элдридж и др., 2000, 2005; Уилер и Бакнер, 2004; Йонелинас и др., 2005; Монтальди и др., 2006), в правильных выражениях. по сравнению с неправильным поиском контекста кодирования (Weis et al. 2004; Tendolkar et al. 2008; Duarte et al. 2011; см. Staresina et al. 2012b для связанного исследования внутричерепной электроэнцефалографии [iEEG]) и в ассоциативном по сравнению с неассоциативным распознаванием (Кирван и Старк, 2004) или распознавание сложных слов (Форд и др.2010). Недавние исследования предполагают, что активность гиппокампа не связана с субъективным чувством воспоминания, а модулируется объемом фактически извлеченной контекстуальной информации (Rugg et al. 2012; Yu et al. 2012).
2012; Yu et al. 2012).
Согласно альтернативной точке зрения, гиппокамп не участвует преимущественно в воспоминании, а не в ознакомлении при контроле силы памяти во время теста на припоминание (Wais 2008; Wais et al. 2010; Smith et al. 2011; см. Montaldi et al. , 2006 для противоположных результатов).Викстед и его коллеги предполагают, что гиппокамп и периринальная кора участвуют как в знакомстве, так и в воспоминании (Squire et al. 2007), но что гиппокамп поддерживает сильные воспоминания, тогда как околоносовая кора поддерживает слабые воспоминания (см. Wais 2008 для метаанализа). Однако они предполагают, что сила памяти сама по себе не является параметром, который отличает гиппокамп от окружающих структур, а скорее то, что различные структуры MTL обрабатывают атрибуты памяти, которые по-разному выражены в сильных и слабых воспоминаниях.Согласно этой точке зрения, гиппокамп поддерживает распознавание многоатрибутных стимулов как на основе воспоминаний, так и на основе знакомства, и его участие в извлечении наиболее очевидно для сильных воспоминаний (Wixted and Squire 2011). Несмотря на различные интерпретации в отношении воспоминаний/знакомств, в целом гиппокамп, по-видимому, в первую очередь участвует в извлечении сильных, богатых, многоатрибутных воспоминаний, тогда как окружающая кора может поддерживать извлечение более простых воспоминаний без участия гиппокампа.
Несмотря на различные интерпретации в отношении воспоминаний/знакомств, в целом гиппокамп, по-видимому, в первую очередь участвует в извлечении сильных, богатых, многоатрибутных воспоминаний, тогда как окружающая кора может поддерживать извлечение более простых воспоминаний без участия гиппокампа.
Воспоминание/знакомство часто используются в литературе о людях для описания как поведенческих феноменов, так и паттернов мозговой активности, коррелирующих с этими феноменами, хотя между двумя типами показателей не обязательно может существовать однозначное соответствие (Voss и Паллер 2010). Ключевым примером является обнаружение того, что околоносовая кора участвует в ассоциативном распознавании памяти в условиях объединения, при котором связанные элементы составляют единый элемент (Diana et al.2010 г.; Форд и др. 2010), тогда как гиппокамп преимущественно участвует в распознавании неунифицированных пар по сравнению с унифицированными (Quamme et al. 2007; Ford et al. 2010). Это было истолковано либо как доказательство того, что знакомство может поддерживать ассоциативное узнавание, когда парные ассоциации могут быть связаны в составную единицу (Ford et al. 2010; Ranganath 2010), либо как свидетельство того, что околоносовая кора участвует в воспоминании объединенных пар (Diana и др., 2010; Викстед и Сквайр, 2011). Точно так же обнаружение нейронов гиппокампа, чувствительных к новизне/старости изображения (Fried et al.1997 год; Рутисхаузер и др. 2006, 2008; Висконтас и др. 2006) подвергались различным интерпретациям, поскольку неясно, как чувствительность нейронов к знакомству соотносится с знакомством на поведенческом уровне (Parks and Yonelinas 2007; Wixted 2007).
2010; Ranganath 2010), либо как свидетельство того, что околоносовая кора участвует в воспоминании объединенных пар (Diana и др., 2010; Викстед и Сквайр, 2011). Точно так же обнаружение нейронов гиппокампа, чувствительных к новизне/старости изображения (Fried et al.1997 год; Рутисхаузер и др. 2006, 2008; Висконтас и др. 2006) подвергались различным интерпретациям, поскольку неясно, как чувствительность нейронов к знакомству соотносится с знакомством на поведенческом уровне (Parks and Yonelinas 2007; Wixted 2007).
Попытки разграничить роль гиппокампа в воспоминании и знакомстве основываются на предположении, что гиппокамп поддерживает устоявшуюся поведенческую диссоциацию между этими двумя проявлениями поиска. Тем не менее, все чаще появляется мнение, что поведенческое различие между воспоминанием и знакомством не отражает основную вычислительную роль (роли) гиппокампа и окружающей его коры (Diana et al.2010 г.; Ранганатх 2010; Восс и Паллер, 2010 г.; Викстед и Сквайр, 2011 г .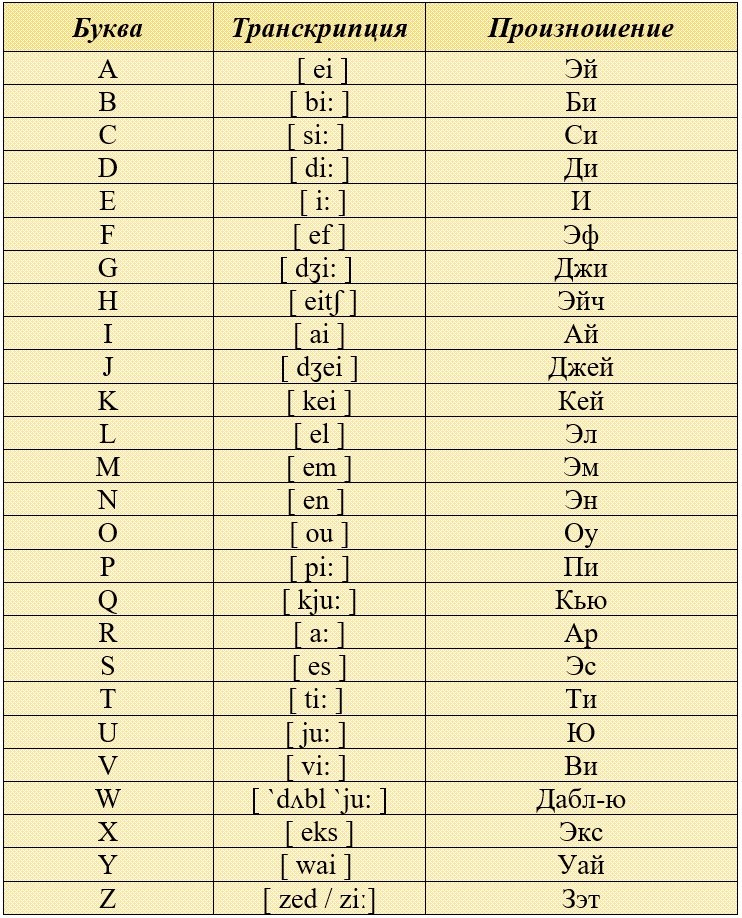 ; Рагг и др. 2012). Альтернативный подход к интерпретации результатов, рассмотренных выше, заключается в том, что формирование гиппокампа играет роль в элементах связывания в их контексте (Diana et al. 2007; Eichenbaum et al. 2007; Ranganath 2010). Эта постулируемая роль также вводит функциональное различие между околоносовой и парагиппокампальной корой, согласно которому околоносовая кора кодирует информацию об элементе, парагиппокампальная кора кодирует контекстную информацию, а гиппокамп кодирует ассоциации предмет-контекст, а также ассоциации предмет-элемент.Сходная точка зрения (Montaldi and Mayes 2010) отводит периринальной коре роль в ассоциации между элементами и элементами памяти, а также внутри домена. Все эти модели подчеркивают роль гиппокампа в поиске связанных ассоциаций, связанных с распознаванием отдельных элементов/контекста, которые поддерживаются окружающей корой. Их также можно рассматривать в соответствии с точкой зрения, упомянутой выше при обсуждении работы с грызунами, что гиппокамп служит картой или указателем, который задействует соответствующие корковые цепи.
; Рагг и др. 2012). Альтернативный подход к интерпретации результатов, рассмотренных выше, заключается в том, что формирование гиппокампа играет роль в элементах связывания в их контексте (Diana et al. 2007; Eichenbaum et al. 2007; Ranganath 2010). Эта постулируемая роль также вводит функциональное различие между околоносовой и парагиппокампальной корой, согласно которому околоносовая кора кодирует информацию об элементе, парагиппокампальная кора кодирует контекстную информацию, а гиппокамп кодирует ассоциации предмет-контекст, а также ассоциации предмет-элемент.Сходная точка зрения (Montaldi and Mayes 2010) отводит периринальной коре роль в ассоциации между элементами и элементами памяти, а также внутри домена. Все эти модели подчеркивают роль гиппокампа в поиске связанных ассоциаций, связанных с распознаванием отдельных элементов/контекста, которые поддерживаются окружающей корой. Их также можно рассматривать в соответствии с точкой зрения, упомянутой выше при обсуждении работы с грызунами, что гиппокамп служит картой или указателем, который задействует соответствующие корковые цепи.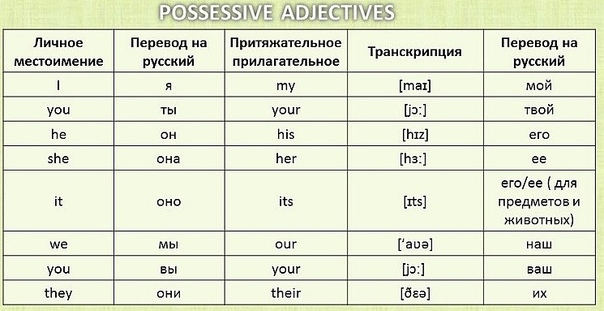
Вышеупомянутые модели предсказывают роль периринальной коры в распознавании отдельных элементов. Это подтверждается исследованиями фМРТ, которые обнаруживают снижение околоносовой активности для очень знакомых раздражителей (Weis et al. 2004; Gonsalves et al. 2005; Daselaar et al. 2006a; Montaldi et al. 2006; хотя см. Kirwan and Stark 2004 об увеличении околоносовой активности). в ответ на правильное ассоциативное распознавание), а также исследование внутричерепной записи, которое обнаружило снижение возбуждения в ответ на знакомые образы в околоносовой области коры (Viskontas et al.2006). Исследование, объединяющее иЭЭГ у пациентов с фМРТ в здоровой группе контроля (Старезина и др., 2012b), рисует более сложную картину. В этом исследовании околоносовая кора продемонстрировала как эффект новизны раннего элемента (дифференциальная реакция на новые и знакомые слова, потенциально отражающий процесс знакомства), так и устойчивый эффект поиска источника (дифференциальная реакция на правильный и неправильный поиск контекста). со словом). Гиппокамп продемонстрировал эффект раннего поиска источника, за которым последовал эффект новизны позднего элемента (потенциально лежащий в основе кодирования нового элемента).Эти исследования показывают четкую роль периринальной коры в распознавании знакомых стимулов.
со словом). Гиппокамп продемонстрировал эффект раннего поиска источника, за которым последовал эффект новизны позднего элемента (потенциально лежащий в основе кодирования нового элемента).Эти исследования показывают четкую роль периринальной коры в распознавании знакомых стимулов.
В целом, эти разные взгляды сходятся на преобладающей роли гиппокампа в ассоциативном поиске. В дополнение к функциональному разделению между гиппокампом и окружающей корой также была предложена функциональная диссоциация внутри гиппокампа, при которой передняя часть гиппокампа больше участвует в кодировании, а задняя — в поиске (Lepage et al., 1998; Strange et al.1999 г.; Спаниол и др. 2009 г.; Поппенк и др. 2013; но см. Schacter and Wagner 1999; Людовик и др. 2008). ФМРТ высокого разрешения (Carr et al. 2010) позволяет проводить альтернативное различие между различными подполями гиппокампа. Несколько исследований, доступных на момент написания этой статьи, по-видимому, выявили диссоциацию между процессами кодирования, наблюдаемыми в DG/CA3, и поиском (или воспоминанием) в CA1/subiculum (Zeineh et al. , 2003; Eldridge et al., 2005; Viskontas et al., 2009). ).
, 2003; Eldridge et al., 2005; Viskontas et al., 2009). ).
Сеть «режим по умолчанию»
Набор областей, состоящий из медиальной префронтальной коры (mPFC), задней поясной извилины, нижней теменной дольки, латеральной височной коры и MTL, был идентифицирован как преимущественно активный в ситуациях, когда внешние раздражители отсутствует.Поэтому этот набор регионов был назван сетью «отдыха» или «режима по умолчанию» (Raichle et al. 2001; Buckner et al. 2008; Raichle 2010). Сети по умолчанию приписывают несколько функций, в том числе независимые от стимулов психические процессы («блуждающие мысли»; Mason et al., 2007), построение мысленных сцен (Hassabis and Maguire, 2007) и самопроекцию в будущее/прошлое (Addis et al. и др., 2007; Бакнер и Кэрролл, 2007).
Существует замечательное перекрытие между сетью, активируемой при извлечении автобиографических воспоминаний (Svoboda et al.2006 г.; Макдермотт и др. 2009) и сеть по умолчанию (Buckner et al. 2008). Кроме того, недавнее исследование пациентов с очаговыми поражениями в регионах в сети по умолчанию показало корреляцию повреждения этой сети с нарушением извлечения автобиографической памяти (Philippi et al. , 2015). В то время как дефицит семантических элементов автобиографической памяти был связан с повреждением левых mPFC и MTL, дефицит эпизодической автобиографической памяти был связан с повреждением правых mPFC и MTL.Метаанализ (Spreng et al., 2009), в котором сравнивались области режима по умолчанию с активациями автобиографического извлечения памяти, проспекта (воображения себя в будущем), навигации и интенциональности («теория разума»), обнаружил значительное совпадение между большинством различные функции. Задачи поиска, поиска и теории разума активировали аналогичные области режима по умолчанию, включая несколько префронтальных, теменных и височных областей. Обилие свидетельств, связывающих эпизодическое припоминание с мысленным построением или моделированием будущих/вымышленных событий (Hassabis and Maguire, 2007, 2009; Hassabis et al.2007 г.; Шактер и Аддис, 2007 г.; Шактер и др. 2008 г.; Бакнер 2010; Рабин и др. 2010 г.; Шпунар 2010; Виард и др. 2012) привело к новому акценту на конструктивном характере памяти (Bartlett 1932).
, 2015). В то время как дефицит семантических элементов автобиографической памяти был связан с повреждением левых mPFC и MTL, дефицит эпизодической автобиографической памяти был связан с повреждением правых mPFC и MTL.Метаанализ (Spreng et al., 2009), в котором сравнивались области режима по умолчанию с активациями автобиографического извлечения памяти, проспекта (воображения себя в будущем), навигации и интенциональности («теория разума»), обнаружил значительное совпадение между большинством различные функции. Задачи поиска, поиска и теории разума активировали аналогичные области режима по умолчанию, включая несколько префронтальных, теменных и височных областей. Обилие свидетельств, связывающих эпизодическое припоминание с мысленным построением или моделированием будущих/вымышленных событий (Hassabis and Maguire, 2007, 2009; Hassabis et al.2007 г.; Шактер и Аддис, 2007 г.; Шактер и др. 2008 г.; Бакнер 2010; Рабин и др. 2010 г.; Шпунар 2010; Виард и др. 2012) привело к новому акценту на конструктивном характере памяти (Bartlett 1932).
Исследование общих черт и различий между эпизодическим воспроизведением во времени, представляющим собой мысленное путешествие во времени в прошлое, и мысленным путешествием во времени в будущее (воображение), позволило отделить чисто мнемонические компоненты от процессов, в более общем плане связанных с построением эпизодов. Аддис и др.2007, 2009; Вейлер и др. 2010 г.; Аддис и Шактер, 2011 г.; Шактер и др. 2012). Примечательно, что гиппокамп, связанный с сетью по умолчанию при поиске (Huijbers et al., 2011), является одной из основных областей, участвующих в задачах, требующих от участника представления будущих сценариев (Addis and Schacter, 2011; Schacter et al., 2012). ; Магуайр и Маллалли, 2013 г.).
Префронтальная кора
Префронтальная кора (ПФК), подобласти которой были упомянуты выше в контексте «сети по умолчанию», была одной из первых областей, связанных с извлечением эпизодической памяти в исследованиях функциональной нейровизуализации (А) (Nyberg et al. др.1995 год; Бакнер и др. 1998 год; Рагг и др. 1998 год; Вагнер и др. 1998 год; Дюзель и др. 1999 г.; Лепаж и др. 2000), в соответствии с исследованиями, выявляющими нарушения памяти у пациентов с повреждением лобных долей (например, Luria, 1966; Stuss et al., 1994; Wheeler et al., 1995; Duarte et al., 2005; Aly et al., 2011). Различные области префронтальной коры участвуют в мнемонических процессах, поддерживающих поиск, особенно в общих процессах предметной области, таких как установка режима поиска или ориентации, инициация попытки поиска, мониторинг, целенаправленное манипулирование полученной информацией и преодоление помех. Shimamura 1995; Rugg et al.1996 год; Москович и Винокур, 2002 г.; Бадре и Вагнер, 2007 г.; Блюменфельд и Ранганат, 2007). Было предложено несколько функциональных подразделений префронтальной коры, исходя из исходной модели асимметрии полушарного кодирования/поиска (HERA), предполагавшей преимущественное участие правой префронтальной коры в припоминании (Tulving et al.
др.1995 год; Бакнер и др. 1998 год; Рагг и др. 1998 год; Вагнер и др. 1998 год; Дюзель и др. 1999 г.; Лепаж и др. 2000), в соответствии с исследованиями, выявляющими нарушения памяти у пациентов с повреждением лобных долей (например, Luria, 1966; Stuss et al., 1994; Wheeler et al., 1995; Duarte et al., 2005; Aly et al., 2011). Различные области префронтальной коры участвуют в мнемонических процессах, поддерживающих поиск, особенно в общих процессах предметной области, таких как установка режима поиска или ориентации, инициация попытки поиска, мониторинг, целенаправленное манипулирование полученной информацией и преодоление помех. Shimamura 1995; Rugg et al.1996 год; Москович и Винокур, 2002 г.; Бадре и Вагнер, 2007 г.; Блюменфельд и Ранганат, 2007). Было предложено несколько функциональных подразделений префронтальной коры, исходя из исходной модели асимметрии полушарного кодирования/поиска (HERA), предполагавшей преимущественное участие правой префронтальной коры в припоминании (Tulving et al. , 1994; Habib et al., 2003; см. Cabeza et al., 2003). альтернативная гипотеза относительно роли левой/правой префронтальной коры в поиске), на внутриполушарные подразделения в соответствии с медиальной/латеральной, передней/задней и дорсальной/вентральной осями, приписывающими определенные мнемонические процессы каждой подобласти (Fletcher and Henson 2001; Wagner et al. .2001 г.; Петридес 2002; Саймонс и др. 2005 г.; Бадре и Вагнер, 2007).
, 1994; Habib et al., 2003; см. Cabeza et al., 2003). альтернативная гипотеза относительно роли левой/правой префронтальной коры в поиске), на внутриполушарные подразделения в соответствии с медиальной/латеральной, передней/задней и дорсальной/вентральной осями, приписывающими определенные мнемонические процессы каждой подобласти (Fletcher and Henson 2001; Wagner et al. .2001 г.; Петридес 2002; Саймонс и др. 2005 г.; Бадре и Вагнер, 2007).
Мозговые корреляты восстановления декларативной памяти человека. ( A ) Мозговая сеть, связанная с памятью распознавания. На рисунке показан метаанализ областей, которые показывают старую> новую активацию в тестах распознавания в 38 исследованиях фМРТ (функциональной магнитно-резонансной томографии). Идентифицированные области включают угловую извилину, хвостатое ядро, дорсолатеральную префронтальную кору (DLPFC), дорсомедиальную префронтальную кору, дорсальную заднюю теменную кору, заднюю поясную кору и предклинье.(На основе данных Кима, 2013 г. ; с разрешения автора.) ( B ) Диаграммы, изображающие динамику быстрой функциональной связи мозговой сети при извлечении памяти, выявленной с помощью электрокортикографической (ЭКоГ) записи у пациентов, подвергающихся мониторингу судорог. Пациенты занимались извлечением пространственных и временных эпизодических контекстов. Фазовая синхронизация между областями мозга использовалась как мера связности. Панели отображают связность, коррелированную с правильным пространственным и временным поиском в диапазонах 1–4 Гц и 7–10 Гц.PHG, парагиппокампальная извилина; MFG, средняя лобная извилина; SFG, верхняя лобная извилина; IFG, нижняя лобная извилина; IPL, нижняя теменная долька; ПКН, предклинье; SPL, верхняя теменная долька. Успешный поиск был связан с большей глобальной связностью между сайтами с медиальной височной долей (MTL), выступающей в качестве центра взаимодействий, но в то время как правильный поиск пространственного контекста характеризовался более низкой частотой взаимодействий в сети, поиск временного контекста характеризовался более быстрым частотные взаимодействия.
; с разрешения автора.) ( B ) Диаграммы, изображающие динамику быстрой функциональной связи мозговой сети при извлечении памяти, выявленной с помощью электрокортикографической (ЭКоГ) записи у пациентов, подвергающихся мониторингу судорог. Пациенты занимались извлечением пространственных и временных эпизодических контекстов. Фазовая синхронизация между областями мозга использовалась как мера связности. Панели отображают связность, коррелированную с правильным пространственным и временным поиском в диапазонах 1–4 Гц и 7–10 Гц.PHG, парагиппокампальная извилина; MFG, средняя лобная извилина; SFG, верхняя лобная извилина; IFG, нижняя лобная извилина; IPL, нижняя теменная долька; ПКН, предклинье; SPL, верхняя теменная долька. Успешный поиск был связан с большей глобальной связностью между сайтами с медиальной височной долей (MTL), выступающей в качестве центра взаимодействий, но в то время как правильный поиск пространственного контекста характеризовался более низкой частотой взаимодействий в сети, поиск временного контекста характеризовался более быстрым частотные взаимодействия. Эти результаты дают представление о том, как несколько контекстов, связанных с одним событием, могут быть получены в одной и той же сети. (На основе данных Watrous et al., 2013; с разрешения авторов.)
Эти результаты дают представление о том, как несколько контекстов, связанных с одним событием, могут быть получены в одной и той же сети. (На основе данных Watrous et al., 2013; с разрешения авторов.)
Метаанализ, сравнивающий кодирование и воспоминание, показал, что как кодирование, так и извлечение меморандумов, таких как изображения и слова, в первую очередь активируют левую префронтальную кору, в отличие от модель HERA (Спаниол и др., 2009). Сравнение объективного и субъективного воспоминаний выявило более сильную активацию объективного воспоминания в вентролатеральной префронтальной коре (VLPFC), дорсолатеральной префронтальной коре (DLPFC) и латеральной передней префронтальной коре, а также более сильную активацию субъективного воспоминания в медиально-передней префронтальной коре, в соответствии с предполагаемой ролью латеральной префронтальной коры в когнитивных процессах. контроль памяти (Simons and Spiers, 2003; Badre and Wagner, 2007) и медиальной префронтальной коры при самореферентной обработке (Northoff et al. 2006).
2006).
Задняя теменная кора
В отличие от результатов функциональной нейровизуализации, касающихся MTL и PFC, существует поразительное несоответствие между данными о поражении задней теменной коры при извлечении и исследованиями пациентов с теменными поражениями. Функциональные нейровизуализационные исследования последовательно наблюдали теменную активацию в эпизодических задачах на припоминание (Shannon and Buckner 2004; Wagner et al. 2005; Vilberg and Rugg 2008; Hutchinson et al. 2009; Spaniol et al. 2009), при этом вентральная задняя теменная кора вовлекалась в первую очередь в воспоминания, а дорсально-задняя теменная часть более широко участвует в узнавании (Shannon and Buckner 2004; Wagner et al.2005 г.; Вилберг и Рагг, 2007, 2008, 2012). И наоборот, в большинстве исследований поражений обнаруживаются ограниченные, если таковые имеются, нарушения при эпизодическом поиске (Berryhill et al., 2007, 2010; Haramati et al., 2008), что подтверждается исследованием повторяющейся транскраниальной магнитной стимуляции (rTMS), которое выявило нарушение задней теменной коры. существенно не ухудшает память (Росси и др., 2006). Исследование, непосредственно сравнивающее пациентов с повреждениями в этой коре головного мозга с фМРТ здоровых людей, выявило анатомическое совпадение между теменной активацией, связанной с воспоминанием, и очагом повреждения у пациентов, которые демонстрировали нормальную производительность при выполнении той же задачи (Simons et al.2008). Сильное участие теменной области в неповрежденном воспоминании в сочетании с относительно тонкими нарушениями, вызванными повреждением этой коры, привело к созданию нескольких моделей, касающихся ее роли в памяти, особенно во внимании к памяти (Cabeza et al. 2008, 2012; Ciaramelli et al. 2008). ), в качестве кратковременного эпизодического буфера (Vilberg and Rugg 2008; Wagner et al. 2005) и в связывающей реляционной активности (Shimamura 2011).
существенно не ухудшает память (Росси и др., 2006). Исследование, непосредственно сравнивающее пациентов с повреждениями в этой коре головного мозга с фМРТ здоровых людей, выявило анатомическое совпадение между теменной активацией, связанной с воспоминанием, и очагом повреждения у пациентов, которые демонстрировали нормальную производительность при выполнении той же задачи (Simons et al.2008). Сильное участие теменной области в неповрежденном воспоминании в сочетании с относительно тонкими нарушениями, вызванными повреждением этой коры, привело к созданию нескольких моделей, касающихся ее роли в памяти, особенно во внимании к памяти (Cabeza et al. 2008, 2012; Ciaramelli et al. 2008). ), в качестве кратковременного эпизодического буфера (Vilberg and Rugg 2008; Wagner et al. 2005) и в связывающей реляционной активности (Shimamura 2011).
Дополнительный вклад в поисковую сеть
До сих пор мы сосредоточились на области, наиболее часто идентифицируемой с извлечением эпизодической памяти человека, однако дополнительные области мозга также были вовлечены в эпизодическое извлечение. Таламусу, в частности передним таламическим ядрам, уделялось особое внимание из-за их связи с гиппокампом. Aggleton and Brown (1999) предположили, что передние таламические ядра и сосцевидные тела необходимы для воспоминаний, тогда как медиальные дорсальные ядра поддерживают знакомство. Недавний обзор (Aggleton et al. 2011) лишь частично поддерживает это предложение. Было показано, что повреждение передних ядер таламуса, которые связаны с гиппокампом и сосцевидными телами, ухудшает опознавательную память, хотя это нарушение значительно менее серьезно, чем повреждение гиппокампа или сосцевидных тел.Исследование фМРТ выявило линейное увеличение активности дорсомедиального таламуса в ответ на изображения, вызывающие более сильное чувство знакомства (Montaldi et al. 2006), однако неясно, ухудшает ли непосредственно повреждение этого ядра распознавание. Дополнительным регионом, к которому возобновился интерес, является ретросплениальная кора (см. также исследования на грызунах выше). Хотя ретросплениальная кора постоянно активируется в исследованиях автобиографического поиска, а повреждение этой области ухудшает восстановление недавних эпизодических воспоминаний, первоначально считалось, что ее роль в памяти непрямая (Vann et al.
Таламусу, в частности передним таламическим ядрам, уделялось особое внимание из-за их связи с гиппокампом. Aggleton and Brown (1999) предположили, что передние таламические ядра и сосцевидные тела необходимы для воспоминаний, тогда как медиальные дорсальные ядра поддерживают знакомство. Недавний обзор (Aggleton et al. 2011) лишь частично поддерживает это предложение. Было показано, что повреждение передних ядер таламуса, которые связаны с гиппокампом и сосцевидными телами, ухудшает опознавательную память, хотя это нарушение значительно менее серьезно, чем повреждение гиппокампа или сосцевидных тел.Исследование фМРТ выявило линейное увеличение активности дорсомедиального таламуса в ответ на изображения, вызывающие более сильное чувство знакомства (Montaldi et al. 2006), однако неясно, ухудшает ли непосредственно повреждение этого ядра распознавание. Дополнительным регионом, к которому возобновился интерес, является ретросплениальная кора (см. также исследования на грызунах выше). Хотя ретросплениальная кора постоянно активируется в исследованиях автобиографического поиска, а повреждение этой области ухудшает восстановление недавних эпизодических воспоминаний, первоначально считалось, что ее роль в памяти непрямая (Vann et al. 2009а). В частности, эта кора преимущественно участвует в воспоминании недавних (Gilboa et al. 2004) и самозначимых воспоминаний (Summerfield et al. 2009), и было высказано предположение, что она поддерживает перевод с эгоцентрических взглядов на аллоцентрические (Byrne et al. 2007). ). Взятые вместе, эти данные указывают на роль ретроспленальной коры в семантизации эпизодических воспоминаний, что может повлечь за собой переход к более аллоцентрическому представлению воспоминаний. Полосатое тело, традиционно связанное с недекларативной памятью, также стало предметом исследований в контексте извлечения информации из декларативной памяти, и было предложено поддерживать когнитивный контроль эпизодического воспроизведения совместно с префронтальной корой (Scimeca and Badre 2012).
2009а). В частности, эта кора преимущественно участвует в воспоминании недавних (Gilboa et al. 2004) и самозначимых воспоминаний (Summerfield et al. 2009), и было высказано предположение, что она поддерживает перевод с эгоцентрических взглядов на аллоцентрические (Byrne et al. 2007). ). Взятые вместе, эти данные указывают на роль ретроспленальной коры в семантизации эпизодических воспоминаний, что может повлечь за собой переход к более аллоцентрическому представлению воспоминаний. Полосатое тело, традиционно связанное с недекларативной памятью, также стало предметом исследований в контексте извлечения информации из декларативной памяти, и было предложено поддерживать когнитивный контроль эпизодического воспроизведения совместно с префронтальной корой (Scimeca and Badre 2012).
Cabeza and Moscovitch (2013) предполагают, что каждая задача памяти задействует различную комбинацию компонентов обработки, некоторые из которых, вероятно, будут общими и для задач, не связанных с памятью. Набор компонентов обработки, которые взаимодействуют для поддержки данной задачи, например, извлечение эпизодической памяти, обозначается как «зависимый от процесса альянс», подчеркивая влияние временных межрегиональных взаимодействий на вклад отдельных областей мозга в выполнение задачи в момент времени. рука. В контексте нашего обсуждения это подчеркивает важность понимания не только роли каждой области мозга в припоминании, но и того, как различные области взаимодействуют для поддержки различных мнемонических функций, включая припоминание (Simons and Spiers, 2003; Mendelsohn et al.2010 г.; Кабеза и др. 2011 г.; Хейберс и др. 2011 г.; Сен-Жак и др. 2011 г.; Форнито и др. 2012 г.; Уотрус и др. 2013). Кроме того, представление о том, что одна и та же область может также участвовать в немемических задачах, может иногда стирать различие между мозговыми сигнатурами восприятия и воспроизведения, возможность, намекаемая выше при обсуждении недавних данных об искусственной активации инграмм-кандидатов в мозгу грызунов. .
рука. В контексте нашего обсуждения это подчеркивает важность понимания не только роли каждой области мозга в припоминании, но и того, как различные области взаимодействуют для поддержки различных мнемонических функций, включая припоминание (Simons and Spiers, 2003; Mendelsohn et al.2010 г.; Кабеза и др. 2011 г.; Хейберс и др. 2011 г.; Сен-Жак и др. 2011 г.; Форнито и др. 2012 г.; Уотрус и др. 2013). Кроме того, представление о том, что одна и та же область может также участвовать в немемических задачах, может иногда стирать различие между мозговыми сигнатурами восприятия и воспроизведения, возможность, намекаемая выше при обсуждении недавних данных об искусственной активации инграмм-кандидатов в мозгу грызунов. .
Распределенные взаимодействия при воспроизведении
Исследования фМРТ показали, что во время воспроизведения взаимодействия между дистальными областями мозга модулируются типом воспроизводимой памяти (Maguire et al.2000 г.; Гринберг и др. 2005), возраст памяти (Viard et al. 2010; Söderlund et al. 2012) и ее достоверность (Mendelsohn et al. 2010). Однако фМРТ дает нам только моментальные снимки состояний мозга, усредненные за период времени, намного более длительный, чем тот, в течение которого функционирует механизм поиска (см. выше). Электрофизиологические исследования пациентов, проходящих обследование перед нейрохирургическим вмешательством, привели к более тонкому пространственно-временному диссекции и новому пониманию подробных механизмов поиска в человеческом мозгу (Suthana and Fried 2012).Ярким примером является межрегиональная фазовая синхронизация. Активированные ансамбли нейронов демонстрируют колебательные паттерны активности, и было высказано предположение, что синхронизация в фазе колебаний между различными областями может способствовать коммуникации и взаимодействию между ними (Fries 2005). Кроме того, синхронизация в разных частотных диапазонах соответствует разным пространственным масштабам, при этом синхронизация на более низких частотах связана с взаимодействием между более отдаленными областями (Canolty et al.
2010; Söderlund et al. 2012) и ее достоверность (Mendelsohn et al. 2010). Однако фМРТ дает нам только моментальные снимки состояний мозга, усредненные за период времени, намного более длительный, чем тот, в течение которого функционирует механизм поиска (см. выше). Электрофизиологические исследования пациентов, проходящих обследование перед нейрохирургическим вмешательством, привели к более тонкому пространственно-временному диссекции и новому пониманию подробных механизмов поиска в человеческом мозгу (Suthana and Fried 2012).Ярким примером является межрегиональная фазовая синхронизация. Активированные ансамбли нейронов демонстрируют колебательные паттерны активности, и было высказано предположение, что синхронизация в фазе колебаний между различными областями может способствовать коммуникации и взаимодействию между ними (Fries 2005). Кроме того, синхронизация в разных частотных диапазонах соответствует разным пространственным масштабам, при этом синхронизация на более низких частотах связана с взаимодействием между более отдаленными областями (Canolty et al.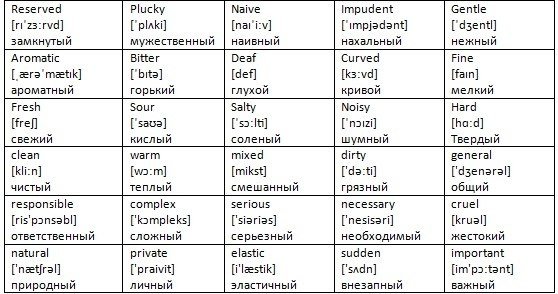 2010). Таким образом, фазовая связь между низкими и высокими частотами может обеспечивать синхронизацию локальной активности в дистальных отделах мозга (Canolty et al., 2006; Jacobs et al., 2007; Canolty and Knight, 2010; Rutishauser et al., 2010; Maris et al., 2011; van дер Мей и др., 2012). Что касается памяти, особый интерес вызывает тета-диапазон (3–8 Гц, включая «медленную тета») и связь тета/гамма (30–150 Гц, включая «высокую гамма»), которые связаны с различными мнемонические процессы кодирования, рабочей памяти и долговременной памяти (Nyhus and Curran 2010; Fell and Axmacher 2011).В частности, успешное восстановление памяти было связано с внутрирегиональным усилением тета- и гамма-колебаний (Осипова и др., 2006; Грубер и др., 2008), обнаруженным в ЭЭГ/МЭГ, а также фазовой синхронизацией между префронтальными, теменными и медиальными височными областями (Андерсон). et al., 2009; Polanía et al., 2012; Watrous et al., 2013) и фазово-амплитудная связь между префронтальными тета- и теменными гамма-колебаниями (Köster et al.
2010). Таким образом, фазовая связь между низкими и высокими частотами может обеспечивать синхронизацию локальной активности в дистальных отделах мозга (Canolty et al., 2006; Jacobs et al., 2007; Canolty and Knight, 2010; Rutishauser et al., 2010; Maris et al., 2011; van дер Мей и др., 2012). Что касается памяти, особый интерес вызывает тета-диапазон (3–8 Гц, включая «медленную тета») и связь тета/гамма (30–150 Гц, включая «высокую гамма»), которые связаны с различными мнемонические процессы кодирования, рабочей памяти и долговременной памяти (Nyhus and Curran 2010; Fell and Axmacher 2011).В частности, успешное восстановление памяти было связано с внутрирегиональным усилением тета- и гамма-колебаний (Осипова и др., 2006; Грубер и др., 2008), обнаруженным в ЭЭГ/МЭГ, а также фазовой синхронизацией между префронтальными, теменными и медиальными височными областями (Андерсон). et al., 2009; Polanía et al., 2012; Watrous et al., 2013) и фазово-амплитудная связь между префронтальными тета- и теменными гамма-колебаниями (Köster et al. , 2014).
, 2014).
Помимо роли этих колебаний в обеспечении межрегиональной синхронизации, несколько моделей предполагают, что тета-колебания могут служить для кодирования определенных временных и пространственных контекстов (Hasselmo 2012; Watrous et al.2013, 2015; Хассельмо и Стерн, 2014 г.). В одном недавнем исследовании Watrous и коллеги (2013) изучали низкочастотную (1–10 Гц) фазовую синхронизацию в электрокортикографических записях парагиппокампальной извилины, теменной коры и латеральной префронтальной коры во время извлечения пространственной и временной информации о местах, ранее встречавшихся во время виртуальная навигация. Они обнаружили общее увеличение предполагаемой сетевой связи (измеряемой по фазовой синхронизации) во время успешного извлечения, в основном сосредоточенное в парагиппокампальной извилине.При сравнении успешного поиска пространственного и временного контекста оба они характеризовались увеличением сетевого подключения, но правильный поиск пространственного контекста включал более низкочастотные взаимодействия по сравнению с поиском временного контекста (B). Это предполагает механизм, который может обеспечить одновременное представление пространственной и временной контекстуальной информации в одной и той же цепи посредством мультиплексирования фазы/частоты (Watrous et al. 2013, 2015). Кроме того, тета-колебания могут поддерживать быстрое переключение между режимами кодирования и поиска в MTL (Hasselmo et al.2002 г.; Хассельмо и Стерн, 2014 г.). Эта модель также подтверждается доказательствами различных колебательных фаз после учебных предметов по сравнению с тестовыми зондами, хотя и в задаче на рабочую память (Rizzuto et al. 2006).
Это предполагает механизм, который может обеспечить одновременное представление пространственной и временной контекстуальной информации в одной и той же цепи посредством мультиплексирования фазы/частоты (Watrous et al. 2013, 2015). Кроме того, тета-колебания могут поддерживать быстрое переключение между режимами кодирования и поиска в MTL (Hasselmo et al.2002 г.; Хассельмо и Стерн, 2014 г.). Эта модель также подтверждается доказательствами различных колебательных фаз после учебных предметов по сравнению с тестовыми зондами, хотя и в задаче на рабочую память (Rizzuto et al. 2006).
Восстановление процессов кодирования во время поиска
Широко представленный взгляд на восстановление эпизодической памяти, упомянутый выше, также в отношении исследований на грызунах, состоит в том, что восстановление влечет за собой восстановление мозговой активности, которая была вызвана во время кодирования (Damasio 1989; McClelland et al.1995 год; рассмотрен в Buckner and Wheeler 2001; Рагг и др. 2008 г.; Данкер и Андерсон, 2010 г.; Рисман и Вагнер, 2012 г.; Леви и Вагнер, 2013). Влиятельные когнитивные теории, основанные на данных, утверждали, что соответствие между сигналами кодирования и воспроизведения и между уровнями обработки (например, в словесных меморандумах) имеет значение для успешного поиска (Талвинг и Томсон, 1973; Моррис и др., 1977). Это порождает вопрос о том, как репрезентация сохраненной памяти может быть связана с репрезентацией опыта.С появлением фМРТ стало возможным исследовать этот вопрос в человеческом мозге более непосредственно, проверяя, влечет ли за собой восстановление специфических моделей активности, зависящих от уровня оксигенации крови (BOLD), которые возникают во время кодирования в отдельных областях мозга.
2008 г.; Данкер и Андерсон, 2010 г.; Рисман и Вагнер, 2012 г.; Леви и Вагнер, 2013). Влиятельные когнитивные теории, основанные на данных, утверждали, что соответствие между сигналами кодирования и воспроизведения и между уровнями обработки (например, в словесных меморандумах) имеет значение для успешного поиска (Талвинг и Томсон, 1973; Моррис и др., 1977). Это порождает вопрос о том, как репрезентация сохраненной памяти может быть связана с репрезентацией опыта.С появлением фМРТ стало возможным исследовать этот вопрос в человеческом мозге более непосредственно, проверяя, влечет ли за собой восстановление специфических моделей активности, зависящих от уровня оксигенации крови (BOLD), которые возникают во время кодирования в отдельных областях мозга.
Первоначальные исследования показали, что сенсорная кора, активная при кодировании, также активна при воспроизведении (Nyberg et al., 2000, 2001; Wheeler et al., 2000, 2006; Vaidya et al., 2002; Gottfried et al., 2004; Kahn et al., 2004). ; Wheeler and Buckner 2004; Woodruff et al.2005). Было высказано предположение, что эта реактивация связана с субъективным опытом воспоминаний (Wheeler and Buckner, 2004), как для достоверных воспоминаний, так и для ложных тревог (Kahn et al., 2004). Джонсон и Рагг (2007) напрямую сравнили активность памяти, специфичную для задачи, с активностью мозга во время кодирования. Они обнаружили перекрытие между областями мозга, лежащими в основе успешного кодирования для конкретной задачи, и областями, лежащими в основе успешного припоминания слов, закодированных в рамках одной и той же задачи (но не в перекрестных задачах).Этот набор исследований дает убедительные доказательства в пользу восстановления коры, то есть восстановления паттернов активности коры, возникающих при кодировании во время извлечения того же события. Однако эти исследования показывают восстановление только на довольно грубом уровне, что указывает на общее совпадение между областями мозга, участвующими в кодировании, специфичном для содержания, и областями, участвующими в извлечении соответствующих стимулов.
; Wheeler and Buckner 2004; Woodruff et al.2005). Было высказано предположение, что эта реактивация связана с субъективным опытом воспоминаний (Wheeler and Buckner, 2004), как для достоверных воспоминаний, так и для ложных тревог (Kahn et al., 2004). Джонсон и Рагг (2007) напрямую сравнили активность памяти, специфичную для задачи, с активностью мозга во время кодирования. Они обнаружили перекрытие между областями мозга, лежащими в основе успешного кодирования для конкретной задачи, и областями, лежащими в основе успешного припоминания слов, закодированных в рамках одной и той же задачи (но не в перекрестных задачах).Этот набор исследований дает убедительные доказательства в пользу восстановления коры, то есть восстановления паттернов активности коры, возникающих при кодировании во время извлечения того же события. Однако эти исследования показывают восстановление только на довольно грубом уровне, что указывает на общее совпадение между областями мозга, участвующими в кодировании, специфичном для содержания, и областями, участвующими в извлечении соответствующих стимулов. Многомерные подходы к анализу данных фМРТ позволяют проводить более детальные тесты восстановления конкретных паттернов пространственной активности в регионах.Традиционный анализ фМРТ фокусируется на выявлении вокселей (объемных пикселей), которые изменяют свою активность в ответ на определенные задачи или когнитивные состояния, тестируя каждый воксел отдельно. Более современные многомерные подходы, такие как анализ многовоксельных паттернов (Хаксби и др., 2001; Норман и др., 2006) и анализ репрезентативного сходства (Кригескорте и др., 2008), смещают акцент на многовоксельные паттерны активности, позволяя идентифицировать восстановление с более высоким пространственным разрешением.
Многомерные подходы к анализу данных фМРТ позволяют проводить более детальные тесты восстановления конкретных паттернов пространственной активности в регионах.Традиционный анализ фМРТ фокусируется на выявлении вокселей (объемных пикселей), которые изменяют свою активность в ответ на определенные задачи или когнитивные состояния, тестируя каждый воксел отдельно. Более современные многомерные подходы, такие как анализ многовоксельных паттернов (Хаксби и др., 2001; Норман и др., 2006) и анализ репрезентативного сходства (Кригескорте и др., 2008), смещают акцент на многовоксельные паттерны активности, позволяя идентифицировать восстановление с более высоким пространственным разрешением.
В первом многовоксельном анализе эпизодической памяти Polyn et al.(2005) обучили классификатор (то есть алгоритм, который позволяет классифицировать состояния мозга на основе различных паттернов активации) на паттернах активности во время кодирования изображений в трех категориях и смогли предсказать категорию изображений, вспоминаемых в свободном тесте на припоминание. Примечательно, что восстановление паттернов активности во время припоминания происходит в отсутствие субъективных воспоминаний (Johnson et al., 2009), а также в ситуациях конкурирующих воспоминаний (Kuhl et al., 2011, 2012), что позволяет предположить, что восстановление может происходить даже в неявном поиске.Повышенное пространственное разрешение, обеспечиваемое такими многомерными подходами, позволяет исследовать корковое восстановление в зависимости от события. В двух недавних исследованиях (Старезина и др., 2012a; Ричи и др., 2013) удалось выявить такое восстановление для конкретных предметов. Старесина и др. (2012a) исследовали, восстанавливаются ли нейронные репрезентации конкретных предметов, возникающие во время эпизодического кодирования, позже при успешном воспоминании. Они представили участникам пары «слово-сцена», а затем проверили их память, используя слова в качестве подсказок, измеряя активность мозга (с помощью фМРТ) как во время кодирования, так и при воспроизведении.
Примечательно, что восстановление паттернов активности во время припоминания происходит в отсутствие субъективных воспоминаний (Johnson et al., 2009), а также в ситуациях конкурирующих воспоминаний (Kuhl et al., 2011, 2012), что позволяет предположить, что восстановление может происходить даже в неявном поиске.Повышенное пространственное разрешение, обеспечиваемое такими многомерными подходами, позволяет исследовать корковое восстановление в зависимости от события. В двух недавних исследованиях (Старезина и др., 2012a; Ричи и др., 2013) удалось выявить такое восстановление для конкретных предметов. Старесина и др. (2012a) исследовали, восстанавливаются ли нейронные репрезентации конкретных предметов, возникающие во время эпизодического кодирования, позже при успешном воспоминании. Они представили участникам пары «слово-сцена», а затем проверили их память, используя слова в качестве подсказок, измеряя активность мозга (с помощью фМРТ) как во время кодирования, так и при воспроизведении. Парагиппокампальная кора показала повышенное сходство между паттернами активации кодирования и извлечения (сходство кодирования-восстановления) во время успешного припоминания. Эта реактивация наблюдалась только во время успешного припоминания, а не во время узнавания, и была специфичной для события (т. Е. Сходство кодирования и извлечения было обусловлено восстановлением определенных комбинаций слово-сцена, а не самой сценой). Примечательно, что сходство кодирования-восстановления гиппокампа не предсказывало память, но амплитуда ответа гиппокампа коррелировала со сходством кодирования-восстановления парагиппокампа во время извлечения, что позволяет предположить, что оно может опосредовать восстановление, наблюдаемое в парагиппокампе.
Парагиппокампальная кора показала повышенное сходство между паттернами активации кодирования и извлечения (сходство кодирования-восстановления) во время успешного припоминания. Эта реактивация наблюдалась только во время успешного припоминания, а не во время узнавания, и была специфичной для события (т. Е. Сходство кодирования и извлечения было обусловлено восстановлением определенных комбинаций слово-сцена, а не самой сценой). Примечательно, что сходство кодирования-восстановления гиппокампа не предсказывало память, но амплитуда ответа гиппокампа коррелировала со сходством кодирования-восстановления парагиппокампа во время извлечения, что позволяет предположить, что оно может опосредовать восстановление, наблюдаемое в парагиппокампе.
Исследования ЭЭГ также показали перекрытие кодирования и извлечения (Rösler et al., 1995; Gratton et al., 1997; Khader et al., 2005). Учитывая низкое временное разрешение фМРТ, которое не позволяет сравнивать временную динамику активности при кодировании и извлечении с разрешением менее нескольких секунд, многообещающим подходом может быть проверка перекрытия кодирования-восстановления с использованием МЭГ (Jafarpour et al. 2014). ) или комбинируя ЭЭГ и фМРТ (Khader et al. 2007; см. Johnson and Rugg 2007 и Johnson et al.2008 г. для двух исследований с использованием аналогичных протоколов с фМРТ и ЭЭГ соответственно). Электрофизиологические измерения, полученные с помощью глубинных электродов у нейрохирургических пациентов, также используются для получения данных с высоким временным разрешением при исследовании извлечения человека. Например, используя этот подход, Gelbard-Sagiv et al. (2008) показали, что припоминание эпизодов с использованием видеоклипов в качестве меморандумов влечет за собой активацию характерных для эпизодов паттернов активности в гиппокампе.
2014). ) или комбинируя ЭЭГ и фМРТ (Khader et al. 2007; см. Johnson and Rugg 2007 и Johnson et al.2008 г. для двух исследований с использованием аналогичных протоколов с фМРТ и ЭЭГ соответственно). Электрофизиологические измерения, полученные с помощью глубинных электродов у нейрохирургических пациентов, также используются для получения данных с высоким временным разрешением при исследовании извлечения человека. Например, используя этот подход, Gelbard-Sagiv et al. (2008) показали, что припоминание эпизодов с использованием видеоклипов в качестве меморандумов влечет за собой активацию характерных для эпизодов паттернов активности в гиппокампе.
Хотя существуют убедительные доказательства того, что активность мозга, связанная с кодированием, частично восстанавливается во время поиска, перекрывающаяся активность по-прежнему составляет небольшую долю от общей мозговой активности, вызываемой во время кодирования и поиска (Johnson and Rugg 2007; Rugg et al.2008 г.; Даселаар и др. 2009 г. ; Ким и др. 2010). Это может быть вызвано несколькими сопутствующими факторами (Rugg et al. 2008), такими как особенности исходного опыта, которые могут быть не закодированы или закодированы, но не извлечены. Кроме того, учитывая реконструктивный характер восстановления памяти (Лофтус и Палмер, 1974; Шактер и др., 1998), в репрезентацию может быть включена дополнительная информация, не имеющая отношения к исходному событию.
; Ким и др. 2010). Это может быть вызвано несколькими сопутствующими факторами (Rugg et al. 2008), такими как особенности исходного опыта, которые могут быть не закодированы или закодированы, но не извлечены. Кроме того, учитывая реконструктивный характер восстановления памяти (Лофтус и Палмер, 1974; Шактер и др., 1998), в репрезентацию может быть включена дополнительная информация, не имеющая отношения к исходному событию.
Действительно, было показано, что ложные воспоминания вызывают ложные эффекты восстановления, имитирующие эффекты достоверного восстановления (Kahn et al.2004), в соответствии с выводами о том, что большинство областей мозга, участвующих в припоминании, чувствительны к субъективному ощущению припоминания, а не к его достоверности (Mendelsohn et al. 2010; Rissman et al. 2010; Dennis et al. 2012). Интересно, что основные области, которые, как было обнаружено, отличают истинные воспоминания от ложных, находятся в сенсорной коре и MTL (Cabeza et al., 2001; Slotnick and Schacter, 2004; Daselaar et al. , 2006b; Kirwan et al., 2009; Dennis et al., 2012). те же регионы, в которых можно ожидать восстановления.
, 2006b; Kirwan et al., 2009; Dennis et al., 2012). те же регионы, в которых можно ожидать восстановления.
В более общем плане доминирующие модели консолидации систем (см. выше, также Squire et al. 2015) предполагают, что представления памяти со временем меняются. Например, сообщалось, что системная консолидация у людей включает семантизацию эпизодического опыта (Cermak 1984; Conway et al. 1997; Nadel et al. 2007; Conway 2009; Piolino et al. 2009; Furman et al. 2012). Одна из основных дискуссий в этом отношении (см. также обсуждение исследований на грызунах выше) относится к тому, зависит ли только извлечение недавних воспоминаний от гиппокампа (т.г., Сквайр и др. 2001 г.; Сквайр и Бэйли, 2007 г.; Барч и др. 2011), или требуется ли это для извлечения ярких воспоминаний, независимо от возраста памяти (Moscovitch et al. 2005, 2006; Hassabis and Maguire 2007; Rosenbaum et al. 2008; Dudai 2012; Bonnici et al. 2013). Данные исследований нейровизуализации человека показывают, что как яркость, так и возраст памяти влияют на различные аспекты активности гиппокампа и его связь с корковой поисковой сетью (например, Gilboa et al. , 2004; Piolino et al.2009 г.; Виард и др. 2010 г.; Фурман и др. 2012 г.; Харанд и др. 2012 г.; Седерлунд и др. 2012). Два недавних исследования анализа многовоксельных паттернов напрямую исследовали, как течение времени влияет на репрезентацию как в гиппокампе, так и в коре головного мозга, путем декодирования конкретных недавних/отдаленных автобиографических воспоминаний (Bonnici et al. 2012, 2013). Они обнаружили, что и vmPFC, и гиппокамп содержат информацию о недавних и отдаленных воспоминаниях, а более отдаленная информация содержится в vmPFC и заднем гиппокампе.
, 2004; Piolino et al.2009 г.; Виард и др. 2010 г.; Фурман и др. 2012 г.; Харанд и др. 2012 г.; Седерлунд и др. 2012). Два недавних исследования анализа многовоксельных паттернов напрямую исследовали, как течение времени влияет на репрезентацию как в гиппокампе, так и в коре головного мозга, путем декодирования конкретных недавних/отдаленных автобиографических воспоминаний (Bonnici et al. 2012, 2013). Они обнаружили, что и vmPFC, и гиппокамп содержат информацию о недавних и отдаленных воспоминаниях, а более отдаленная информация содержится в vmPFC и заднем гиппокампе.
Репрезентации воспоминаний также могут изменяться при обнаружении в поиске новых событий, связанных с существующими воспоминаниями (например,г., в реконсолидации; Надер 2015). Такие условия могут привести к обобщению/выводу (Moses et al. 2006; Shohamy and Wagner 2008; Kumaran and McClelland 2012), интерференции (Müller and Pilzecker 1900; Anderson 2003; Wixted 2004) или созданию ложных воспоминаний (Schacter and Slotnick 2004). ; Лофтус, 2005). Все более чувствительные методы анализа визуализации человеческого мозга обеспечивают улучшенные возможности для исследования взаимодействия между кодированием новой информации и реактивацией предыдущих представлений (Старезина и др.2012б; Бен-Яков и др. 2014; Браун и др. 2015).
; Лофтус, 2005). Все более чувствительные методы анализа визуализации человеческого мозга обеспечивают улучшенные возможности для исследования взаимодействия между кодированием новой информации и реактивацией предыдущих представлений (Старезина и др.2012б; Бен-Яков и др. 2014; Браун и др. 2015).
В целом, исследования, описанные в этом разделе, показывают, что восстановление эпизодической памяти в человеческом мозгу влечет за собой активацию кортико-гиппокампальных или корковых представительств. Блокировка во времени преходящих пространственно-временных паттернов активации в отдельных распределенных ансамблях коры или кортико-гиппокампа может позволить перестановочное связывание репрезентативных элементов для получения специфических репрезентаций, а также может позволить хранить разные репрезентации в одной и той же сети.Эти представления во многом обусловлены активностью во время кодирования, но они податливы, подвержены влиянию времени, семантизации и взаимодействию существующей памяти с новой информацией.